If you’re a regular TetZoo reader you’ll be familiar with my several articles on the life appearance of Mesozoic dinosaurs, an issue in which I have a special interest. Alas, several of these articles were published at TetZoo ver 2 – the ScienceBlogs years – and hosting issues at the site concerned mean that they’re currently appearing sans all of the many images I so lovingly uploaded. Which is a pain.

A really nice, life-sized model of a titanosaurian sauropod, on display in Romania and constructed by Brian Cooley. Image: Darren Naish.
Partly as a consequence, and partly because the issue is on my mind due to several concurrent projects (he says, cryptically), now is a good time to talk once more about the life appearance of Mesozoic dinosaurs, and I’m going to start with sauropods. Sauropods have been covered quite a few times on TetZoo before, as you can see from the list of links at the bottom of this article.
The head, and face especially. We’ll start with the head. Sauropod skulls were proportionally small compared to the overall size of the animals, but not comically so. It’s also worth saying that their eyes – while pretty big in absolute size (based on the size of the eye socket and the sclerotic rings preserved in fossils) – are easy to over-emphasise in illustrations. If you’ve heard that sauropods might have had trunks and thought it reasonable or plausible… well, it’s a poor idea that’s neither reasonable nor plausible, and has a lot counting against it, as covered in the ver 3 article here.

Did sauropods have trunks? NO. It’s the dumbest idea ever, and every piece of evidence counts against it. Several authors have independently explored this idea, including Robert Bakker (upper right) and Bill Munns (lower right). The two images at left are from Knoll et al.’s (2006) paper refuting the trunk idea. Images: Knoll et al. (2006), Bakker (1986), Bill Munns (original here).
Trunks are a no, but did sauropods have ‘lips’ or ‘cheeks’? A whole article could be written on this issue. For now, I’ll summarise things by saying that sauropod skull bone texture indicates that they likely did have extra-oral tissues that mostly covered their teeth (Witton 2018), meaning that they were facially similar to lizards and kin.
A suggestion that some titanosaurs had blade-like cutting sections in the posterior sections of their jaws – so-called ‘guillotine crests’ (Apesteguía 2004) – looks unlikely given that it would require the animals concerned to do their food procurement at the sides of the jaws, rather than at the front like all other sauropods and other dinosaurs. It’s more likely that a mistake has been made here and that the jaw segments thought to support those blade-like, keratinised sections were, while sharp-edged, covered in normal lip tissue. The possibility that diplodocoids and maybe other sauropods too might have had true beaks at the font of the mouth has recently been put forward, but so far only in preliminary fashion.

Apesteguía (2004) argued that some titanosaurs - this is the rhino-sized Bonitasaura - had keratinised ‘guillotine crests’ on the edges of the jaws. I think that this is likely a mistake. Image: Apesteguía (2004).
The nostrils and nose. As is now widely known, and near-universally accepted, the external or fleshy nostrils of sauropods were almost certainly not located way up on the forehead, far from the front of the snout, as was long thought based on the retracted position of the bony nostril openings. A more ‘normal’, anterior position for the nostrils was demonstrated by Witmer (2001) who pointed to evidence from blood vessel and nerve impressions and associated cranial hollows, all of which are located on the anterior part of the snout. These indicate the most likely position of the fleshy nostril and associated blood vessel clusters.

The soft dinosaur revolution comes to Sauropod Town. At left, an image by Matt Wedel which shows why we need more soft tissue on our sauropods. At right, Larry Witmer’s (2001) depiction of the different possibilities as goes nostril position in sauropods. (a) is the most likely option based on anatomical data. Image: Mathew Wedel, Witmer (2001).
There are also reasons for thinking that the giant, cavernous bony nostril openings and tall nasal bars of some macronarian sauropods supported, and were surrounded by, dome-shaped soft tissue convexities, superficially recalling the bulging nasal regions of some living monitor lizards. Part of my reason for saying this comes from the nasal anatomy of the exquisitely preserved South American titanosaur Sarmientosaurus. Here, an anteriorly projecting bony spine located along the midline and associated bar-like structures on the lateral edges of the large narial fossa – the big bony opening surrounding the bony nostril – indicate that a wide, convex mass of tissue connected the forehead with the sides and front of the snout’s upper surface (Martínez et al. 2016). Sarmientosaurus isn’t unique in this respect but is a particularly good example.

The skulls of some sauropods - this is the titanosaur Sarmientosaurus - indicate that there were bulbous nasal structures covering much of the snout region. Image: WitmerLab.
Indeed, skull openings in general were almost definitely not sunken in appearance or otherwise all that obvious, something that’s being said of archosaurian faces in general as artists and anatomists have learnt to take better attention of the conditions present in living animals (where cranial openings are just about never obvious in the live creature).
Accordingly, sauropod faces were seemingly ‘softer’ and more ‘padded’ than convention would have it, at least some of – perhaps all of – the nasal and forehead region being convex and fleshy, rather than shrink-wrapped and covered in thin skin alone (Witton 2018).
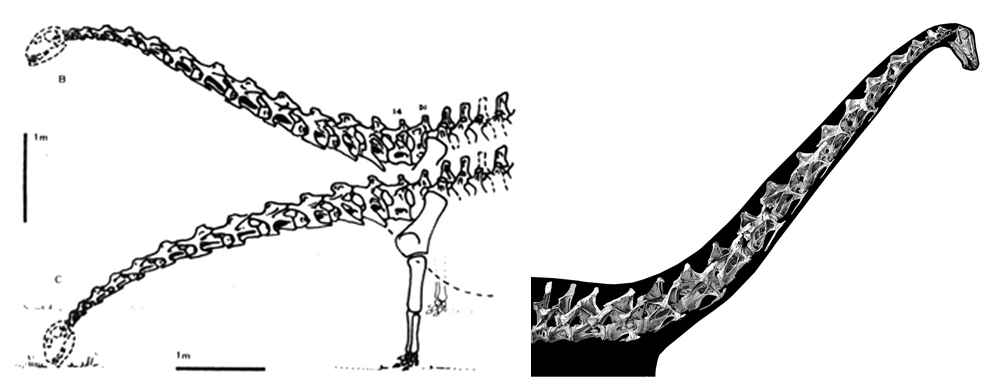
Were sauropod necks mostly semi-horizontal and with a limited range of motion, as argued by Martin (1987) and illustrated at left? Nope; it’s more likely that the necks were often held aloft and far more flexible, as argued by Taylor et al. (2009) and depicted at right. Images: Martin (1987), Taylor et al. (2009).
The neck. The most remarkable feature of sauropod anatomy is the neck. As goes how the neck was held and how flexible it was, several possibilities have been put forward, some workers arguing that it was held in a mostly horizontal attitude for much of the time and with only a limited range of lateral and vertical flexibility. I’m part of a group who argue for mostly elevated habitual neck poses (even in diplodocids) and a wide range of flexibility (Taylor et al. 2009). Arguments that sauropods must have been horizontal-necked do not take account of the flexibility permitted by cartilage, or – in living animals of all sorts – the ranges of motion that happen at zygapophyseal junctions and at the neck base and head-neck junctions.

There are good reasons for thinking that sauropod necks were habitually inclined upwards, as is typical for terrestrial tetrapods in general, and several fossils are actually preserved with the neck in this orientation. This montage by Greg Paul shows several of the fossil concerned. Image: Paul (1998).
It's been convention to show the sauropod neck as a featureless tubular structure, like a hose. This is mostly wrong, in part because the shapes of the vertebrae reveal a more unusual cross-sectional shape that varies from one sauropod group to the next. In some sauropods (like mamenchisaurs), the vertebrae are quite narrow and the neck would have looked laterally compressed in places, as it is (for at least some of its length) in giraffes. In diplodocoids – apatosaurines in particular – the neck is narrow close to the head but wide for much of its length, and subtriangular in cross-section, being widest across the neck’s underside. A neck that was quite narrow in its anteriormost quarter or so but was markedly wide for the rest of its length seems to have been the case in brachiosaurs and at least some titanosaurs.

The necks of some sauropods - a good example is the ultra-long-necked Omeisaurus, photographed at Zigong Dinosaur Museum - are somewhat laterally compressed for at least part of their length. Image: Darren Naish.
Indeed, the very base of the neck is remarkably broad in some sauropods – Camarasaurus is the classic example, where the neck base is not that different in width from the front of the chest – meaning that the neck would taper gradually along its length if you were looking at the animal from the front or back, or from above or below.

The neck bases of some sauropods - this is the Camarasaurus replica on display in London’s NHM - are shockingly broad. It would be wrong to show the neck as a narrow, hose-like object with parallel sides. Image: Darren Naish.
The vertebrae themselves are very complicated with large hollows on their sides, projecting neural spines on the apices and so on. In the most extreme version of the shrink-wrapping meme promoted by one or two palaeontologists and palaeoartists, sauropod necks have been depicted as if these structures should be visible in the live animal, Ely Kish’s apatosaurines from 1983 being the ultimate example. This was definitely not the case. It remains uncertain how much musculature and soft tissue surrounded the vertebrae, but it was almost certainly (based on the anatomy of living animals) enough to obscure the form of the vertebrae, their bulbous junctions perhaps being visible in sauropods of some or many sorts.

The brilliant, late Ely Kish, one of the best palaeoartists of all time, constructing a scale model of Apatosaurus (in preparation for a large painting). Partly on the advice of palaeontologist Dale Russell, she depicted great lateral concavities on the sides of the neck. Image: Russell (1987).
Necks as display banners. The unprecedented form of the sauropod neck makes it plausible – perhaps even likely – that the neck was used as a display structure. Phil Senter’s suggestion that the size and length of the neck was driven, in evolutionary terms, by its use as a display structure (Senter 2007) is not supported by evidence (Taylor et al. 2011). However, this doesn’t discount co-option of the neck in display, so it really is – while wholly speculative right now – worth taking seriously the possibility that display structures of various kinds could well have adorned sauropod necks. These could include wattles, dewlaps, spiky frills, spines, filaments, inflatable pouches or distensible flags or flaps. A few artists have explored these possibilities, most notably Brian Engh and Emiliano Troco.

As weird as it might seem, it is at least plausible that the sauropod neck was decorated with weird soft-tissue display structures, as depicted here on a diplodocid. Image: Emiliano Troco.
Hands. Sauropod hands are extremely odd, and very different from the rounded, elephant-style structures, edged with big nails and/or several claws, shown so often in artistic reconstructions and museum models. Indeed, they’re sufficiently weird and interesting that I’ve written whole articles about them before but, as I said above, these are currently lacking all of their relevant illustrations and are thus all but useless.

Sauropod hands are essentially unique. They’re weird, semi-tubular structures with pillar-like metacarpals. At left, a brachiosaur hand. At right, the hand of the turiasaur Zby. Images: Anthony Maltese, Darren Naish.
The sauropod hand is essentially a semi-tubular structure formed of elongate metacarpals arranged, pillar-like, in a semi-circle. The posterior surface – corresponding to the palm – was hollow, the consequence being a semilunate area of contact with the ground. We’ve known since at least 1940 that sauropod hands had this very unusual form thanks to fossil tracks (Falkingham et al. 2014), which makes it all the weirder that people have ignored this information and merrily continued giving sauropods elephant-like hands across the decades.

Roland T. Bird’s sauropod track illustrations from the 1940s - shown here - clearly show the true, highly unusual form of the sauropod manus. Shame on those who ignored this information in the following decades. Image: Falkingham et al. (2014), CC BY 4.0.
Tracks also show that nails and claws were absent from the hands, except on the thumb where a pointed claw – which varied considerably in exact shape and size from one group to the next – projected inwards and slightly backwards. The thumb claw was lost within Titanosauria, meaning that at least some members of this group lacked nails and claws on their hands altogether. I will concede that poorly defined convexities corresponding to manual digits were present in at least some sauropods, but they still wouldn’t have looked like distinct digits.
Some tracks appear to show that tough semi-conical tubercles projected from the skin on the front and sides of the hand, perhaps giving part of the hand a spiky or tuberculated appearance (Milàn et al. 2005). Maybe these structures had a role in foraging, digging, display or combat, since it’s plausible that they made the hands gnarlier and harder than they would have been otherwise.

Vertical scores associated with sauropod hand prints indicate that at least some of them had tuberculate hand skin, as depicted here (at right) in this modified version of a Greg Paul illustration. Image: Milàn et al. (2005).
Feet. Three large, curved claws projected anterolaterally from the inner three toes. At least some sauropods possessed four such claws. The outer two toes projected as blunt, rounded convexities in some, most or all sauropods. This is obvious from at least some Brontopodus tracks (Meyer et al. 1994). They might have had nails but were more likely lacking horny structures of any sort and essentially continuous with the rest of the foot’s outer surface. In contrast to the hand, the foot did have a massive fatty pad at its back.
Until recently it was thought that the metatarsals were held at a high angle, the consequence being a short foot shaped like that of an elephant but for the claws (Paul 1987). Data from articulated skeletons and tracks, however, show that the metatarsus was not as erect as argued, in which case the toes were longer and flatter than depicted by some artists.

Tschopp et al. (2015) used data from complete Camarasaurus hands and feet to produce the skeletal and soft-tissue models you see here (in 1 and 2), and then used these to generate tracks (3). The tracks are an exact match for real fossil tracks. Image: (c) Tschopp et al. (2015).
On the subject on feet and limbs, trackways also show that sauropods differed in how they placed their hands and feet, some walking with a very narrow gait (the hands and feet being placed close to the midline), others using a wide gait (where there was some short distance between the hands and feet of the left side versus those of the right), and others being intermediate. It should be remembered that even the widest-gauge sauropods did not walk with their feet all that far apart, but that the legs were almost certainly angled inwards, as is typical in living animals. For more on this issue see Scott Hartman’s article here.
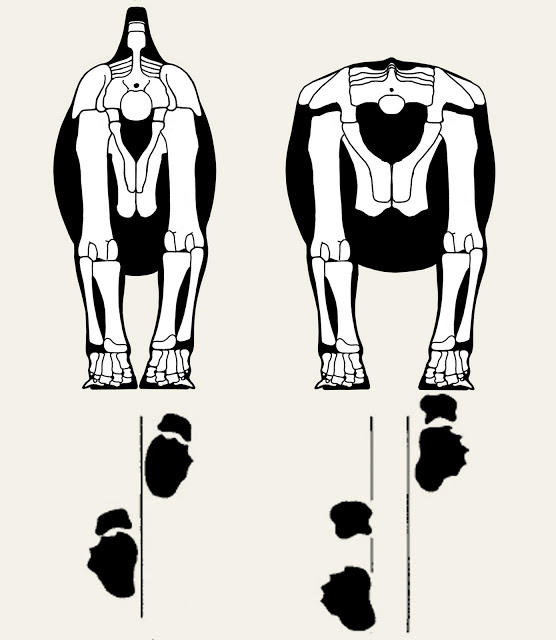
Sauropods were variable in cross-sectional shape and in whether they walked with narrow-gauge or wide-gauge gaits, as depicted here by Scott Hartman. Image: Scott Hartman’s skeletaldrawing.com
The body and tail. Sauropod bodies were variable in cross-sectional shape, length and other details. Diplodocoids, for example, were relatively narrow, deep-bodied and with a tall ridge formed from their neural spines running along the back, while titanosaurs were extremely broad across the hips and must have been just about flat across the back. The thorax may have sloped down ever so slightly in diplodocoids (in part because their forelimbs were shorter than their hindlimbs) whereas the thorax was angled upwards slightly or even markedly in some macronarians, like brachiosaurs and some titanosaurs.
Articulated skeletons show that the tail mostly projected horizontally from the pelvis (a subtle arch at the tail base is present in diplodocoids and some others, in fact), but the macronarians with the upward-sloping bodies also had a downward-sloping tail. A real curiosity is provided by the tails of some mamenchisaurs which seem to have projected upwards at an angle. This has been discussed and illustrated by Paul (2010) and more recently by Hallett & Wedel (2016), but has otherwise gone undiscussed as far as I know. It sounds so odd that surely some mistake has been made… though I really don’t think it has.

Tall neural spines, massive, wing-shaped transverse processes and other structures show that an enormous quantity of musculature would have been obvious along the proximal part of the sauropod tail. This is the tail of … ugh .. Dippy, the cast of Diplodocus carnegii until recently on show in London. Image: Darren Naish.
As is typical for non-bird dinosaurs, the musculature at the base of the tail was (so we can say from the relevant bony attachment points) evidently enormous and bulky, the proximal part of the tail likely being similar in width to the pelvis and thighs combined.
The integument. We know for definite that sauropods of all groups were scaly animals, since scaly skin impressions and actual preserved skin patches are known for diplodocids, Camarasaurus and titanosaurs at least. A few skin folds here and there were almost certainly present (say, where the limbs met the body, at the limb joints, and at mobile zones in the neck) but a fissured, wrinkled or scored elephant-like skin is a big fat no and every effort should be made to avoid it in artistic depictions.

Haestasaurus from the English Wealden (shown at left) and a few other sauropods preserve polygonal scales that vary somewhat in size. Image: Darren Naish, Czerkas (1994).
Polygonal – specifically, pentagonal, hexagonal and heptagonal – scales are known for the macronarian Haestasaurus and some diplodocids, while rounded scales possessing a papilliform texture of tiny bumps are also preserved in a diplodocid specimen (Czerkas 1994). The polygonal structures varied in size somewhat, those present in more mobile parts of the skin (say, the inner crease of the elbow) being smaller than those present in non-mobile sections. Even so, the largest scales were, at most, 60 mm across, which is not large at all on an animal more than 15 m long. This is pretty common in non-bird dinosaurs, by the way. The scales were often so small that they wouldn’t be readily visible from any distance greater than a few metres. The papilliform texture on those diplodocid scales seems to have been widespread across sauropods and means that sauropod skin would have been rough to the touch.
A diplodocid specimen from Wyoming that has scaly skin preserved also preserves tall, conical dermal spines (note: they are not horn-covered bony structures) that must have been arranged along the dorsal midline (Czerkas 1992). These are variable in height (the biggest are 18 cm tall) and seem to have formed a single row along the top of the tail’s proximal part at least. This specimen was originally implied to belong to Diplodocus but is of indeterminate identity. It might belong to Kaatedocus, since remains of this dinosaur come from the exact same quarry.

Large, laterally compressed conical and semi-conical dermal structures lined the upper surface of the tail (at least) in some diplodocids, and perhaps in other diplodocoids and sauropods too. Image: Czerkas (1994).

The conical and semi-conical dermal spines of some diplodocids were variable in height, breadth and shape, as illustrated here. Image: Czerkas (1994).
Were these structures present across all diplodocids, all diplodocoids, all neosauropods or even all sauropods, or were they exclusive to one small clade, perhaps even just to Kaatedocus itself? In the absence of further information we can’t say, but it’s appropriate to depict them on diplodocids and their close kin at least, in the absence of further information. The possibility that shorter and/or taller dermal structures of this sort were present elsewhere on other sauropods exists.

The discovery of those conical and semi-conical spines in a diplodocid led Stephen Czerkas to produce this new look for these dinosaurs. It might be correct, but it is not clear whether the structures were as extensive along the animal’s length as shown here. Image: Czerkas (1992).
Horn-covered osteoderms - that is, lumps and nodules with a bony core - were arranged across the backs and flanks of some titanosaurs, specifically the members of the clade Lithostrotia. These structures were variously rounded or oval, sometimes quite flat and sometimes slightly or strongly domed. They appear to have been numerous and prominent in some of the relevant species and would have given their backs and sides a pebbly, armoured appearance.
Colours. There’s a traditional view stating that big dinosaurs were likely grey and plain because big living mammals are, as is the Komodo dragon, I suppose. I’m going to be bold here and say that there’s no reason whatsoever to take any notice of this. Sauropods were not mammals or Komodo dragons. While an argument could be made that camouflage would have been beneficial, and that certain pigments and patterns would have been helpful or necessary for physiological reasons (heat-shedding, heat retention, UV protection and so on), bold and complex patterns and bright, even vibrant colours are all consistent with the ecophysiological demands of being a sauropod, and the good colour vision and complexity of sauropod integument, and likely reliance on visual display, mean that they very likely could have been more like gigantic lizards or birds than elephants. In any case, giraffes, perenties and others show that big, terrestrial animals don’t have to be bland and grey as has often been stated.

The idea that big terrestrial animals have to be bland never was correct. Perenties Varanus giganteus might not be that colourful, but they help emphasise the possibility that striking patterns can be present even in very large terrestrial reptiles. Image: (c) Stephen Zozaya.
In the absence of any direct evidence, it seems reasonable to me to reconstruct barred, striped, reticulated or dappled colour schemes, to depict bright colours on faces, necks or other areas considered relevant to display, and to imagine any colours reasonable for big, terrestrial animals associated with woodlands, parks, scrubby places, mangroves and all the other habitats frequented by this long-lived, diverse group of animals.

Want to know more about sauropod life appearance? It sometimes surprises people that there are scarcely any good books dedicated to specific Mesozoic dinosaur groups. For sauropods, the main must-have volume is Mark Hallett and Matt Wedel’s The Sauropod Dinosaurs. It’s really good. Dinosaurs Past and Present Vol II is also worth getting, in part because it includes Greg Paul’s (now quite dated, but still useful) article on dinosaur life appearance. Images: amazon (here and here).
If we really want to imagine sauropods as living animals, there is – of course – so much more to say. Body language, posture, gait, social life, anti-predator responses, feeding behaviours, digestive function, sleep and so much more are all things that need to be considered. But that’s where I’ll stop for now. I hope you found this interesting, and we’ll be looking at dinosaur life appearance again sometime soon.
For previous TetZoo articles on sauropods, see…
The hands of sauropods: horseshoes, spiky columns, stumps and banana shapes, October 2008
Sauropod dinosaurs held their necks in high, raised postures, May 2009
Necks for sex? No thank you, we're sauropod dinosaurs, May 2011
The sauropod viviparity meme, May 2011
The Second International Workshop on the Biology of Sauropod Dinosaurs (part I), December 2011
The Second International Workshop on the Biology of Sauropod Dinosaurs (part II), January 2012
Junk in the trunk: why sauropod dinosaurs did not possess trunks, November 2012
The changing life appearance of dinosaurs, September 2014
Brian J. Ford's Aquatic Dinosaurs, 2014 Edition, February 2015
That Brontosaurus Thing, April 2015
10 Long, Happy Years of Xenoposeidon, November 2017
Refs - -
Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften 91, 493-497.
Czerkas, S. A. 1992. Discovery of dermal spines reveals a new look for sauropod dinosaurs. Geology 20, 1068-1070.
Czerkas, S. A. 1994. The history and interpretation of sauropod skin impressions. Gaia 10, 173-182.
Knoll, F., Galton, P. M. & López-Antoñanzas, R. 2006. Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus. Geobios 39, 215-221.
Martin, J. 1987. Mobility and feeding of Cetiosaurus (saurischia, sauropoda [sic]) - why the long neck? In Currie, P. J. & Koster, E. H.(eds) Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Boxtree Books (Drumheller, Alberta), pp. 154-159.
Meyer, C. A., Lockley, M. G., Robinson, J. W. & dos Santos, V. F. 1994. A comparison of well-preserved sauropod tracks from the Late Jurassic of Portugal and the western United States: evidence and implications. Gaia 10, 57-64.
Milàn, J., Christiansen, P. & Mateus, O. 2005. A three-dimensionally preserved sauropod manus impression from the Upper Jurassic of Portugal: implications for sauropod manus shape and locomotor mechanics. Kaupia 14, 47-52.
Paul, G. S. 1987. The science and art of restoring the life appearance of dinosaurs and their relatives - a rigorous how-to guide. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present Vol. II. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and London), pp. 4-49.
Paul, G.S. 1998. Terramegathermy and Cope’s rule in the land of titans. Modern Geology 23, 179-217.
Paul, G. S. 2010. Dinosaurs: A Field Guide. A & C Black, London.
Russell, D. A. 1987. Models and paintings of North American dinosaurs. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present, Volume I. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and Washington), pp. 114-131.
Senter, P. 2007. Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation. Journal of Zoology 271, 45-53.
Taylor, M. P., Hone, D. W. E., Wedel, M. J., & Naish, D. 2011. The long necks of sauropods did not evolve primarily through sexual selection. Journal of Zoology 285, 150-161.
Taylor, M. P., Wedel, M. J. & Naish, D. 2009. Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica 54, 213-220.
Witmer, L. M. 2001. Nostril position in dinosaurs and other vertebrates and its significance for nasal function. Science 293, 850-853.



