Finally, it’s time to talk about the zebras…

Caption: striping is variable.. and that’s a theme you’re about to see discussed a whole lot more. These zebras were photographed in Etosha National Park, Namibia, and are supposed to be Burchell’s zebra. Image: Olga Ernst, CC BY-SA 4.0 (original here).
If you know very little about animals, you might assume that ‘zebras’ are all essentially the same. If you know a fair bit about animals, you likely know that zebras involve three different wild horse lineages: the Plains zebras and Quagga Equus quagga, the long-faced, densely striped Grevy’s zebra E. grevyi, and the odd mountain zebras E. zebra. What I’ve just described is zebra 101. In this article we’re going to go a bit deeper. There’s more to zebra diversity than you might think or know.

Caption: opportunities to see multiple zebra taxa together are rare. Walter Rothschild’s museum at Tring (aka The Natural History Museum at Tring) has a good selection, of which this photo shows part. The sitting zebra at far right is a Burchell’s zebra. Image: Darren Naish.
Zebras are, of course, striped, African members of the horse family (Equidae). But those three zebra lineages are different enough – in proportions, skull shape and pigmentation – that we might doubt the idea that they’re all closely related. Indeed, some studies don’t find zebras to form a clade, but to instead be disparate lineages across the equid tree (Bennett 1980, Orlando et al. 2009). I always liked this idea, but recent genetic work doesn’t support it: zebras apparently are a clade after all (e.g., George & Ryder 1986, Leonard et al. 2005, Steiner & Ryder 2011, Steiner et al. 2012, Vilstrup et al. 2013).

Caption: I very much like illustrations that show the different members of a given group side by side. This image is by Wilhelm Eigener and appeared in Hans Hvass’s 1961 Mammals of the World. At top left is a mountain zebra, and to the right is a Grevy’s zebra. Centre-left is a Chapman’s zebra, with a Burchell’s zebra to the right. At bottom, we see a Grant’s zebra and (in the foregound) an animal that the book calls Boehm’s zebra. Image: Wilhelm Eigener/Hvass (1961).
Today it’s quite well known that DNA has shown the recently extinct Quagga (the last captive individual of which died in Amsterdam Zoo in 1883) to be sufficiently closely related to plains zebras that all should be lumped into the same species (Rau 1978, Higuchi et al. 1984, 1987, Groves & Bell 2004, Leonard et al. 2005, Groves & Grubb 2011, Caro et al. 2014, Larison et al. 2015), the oldest name for which is Equus quagga Boddaert, 1785. Exactly what the Quagga is and how it relates to other equids is a historically complex issue that I must avoid here. I might return to it in time, but for now all you need to know is that the Quagga is nested within the species known technically as Equus quagga, and vernacularly as the Plains zebra.
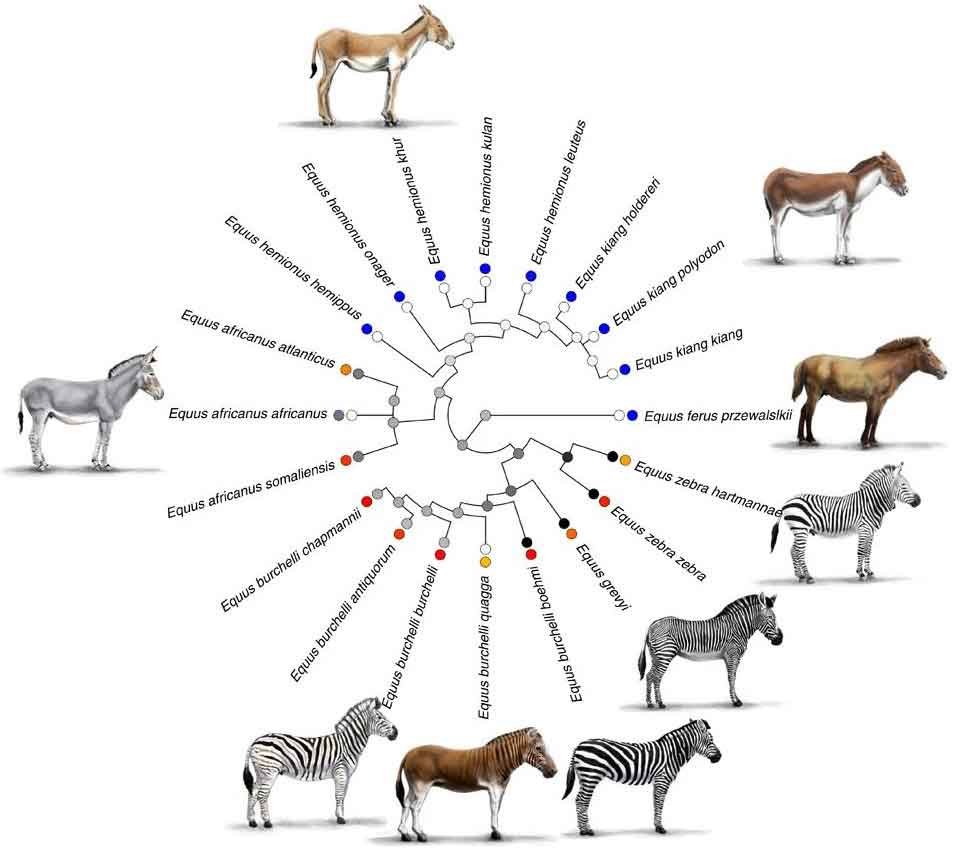
Caption: an equid cladogram, showing current thinking on how the Quagga is nested within the plains zebra. This diagram is from Caro et al. (2014), a paper that focuses on how biting fly density correlates with stripe presence in equids (this explains the coloured circles; I’ve removed the key as it’s a distraction). Equid illustrations by Rickesh Patel.
The term quagga – ‘kwagga’ in Afrikaans and taken from a San word – was originally applied to all zebras and only later (I am of course speaking of the English language alone) restricted to the brown-bodied animals of the south (Barnaby 1996). Zebra – originally applied among African animals to mountain zebras of the south, and apparently based on the portuguese word ‘zevro’* – entered the English language in the 1600s and was first used for animals regarded as interesting quaggas. Much could be written on this issue; this is just a summary. The term dauw was also used for zebras in the 1800s. Its origins are opaque, but some sources say it comes from an indigenous African language.
* An entire article could be written on the idea that ‘zevro’ comes from ‘zebro’, and that this was originally used for the recently extinct Iberian E. hydruntinus. According to this idea, the ‘original zebro/zebra’ was thus a distinct European wild ass. The latest on the claimed zebro/E. hydruntinus link is that E. hydruntinus probably did not survive into modern times, and thus was likely not the ‘original zebro/zebra’ after all (Crees & Turvey 2014).

Caption: two equid-themed books that I’ve used a lot when looking for information on zebras.
The variable Quagga. If you asked me to describe a Quagga, I would say that it’s a ‘reduced-striped’ zebra where the body is dark brown, the legs lack stripes, and striping is limited to the head and neck. That’s not a bad generalisation, but it’s not an accurate one when we look at diversity across known Quagga specimens.
Caption: the only Quagga photographed alive was the London Zoo mare; several photos exist, and this is the best known of them. It was taken in 1870. Image in public domain.
We have 23 preserved Quagga skins, and while some (like the Darmstadt and Amsterdam specimens) have mostly plain, dark brown bodies, most have obvious body striping and some (the Frankfurt specimen in particular) have body striping about as prominent as that of other plains zebras. The Tring Quagga is notable in that its body striping is not just prominent and obvious, but chaotic and ‘zig-zaggy’, with numerous shadow stripes and poor differentiation between striped and stripe-less zones.

Caption: the Tring Quagga shows that at least some Quaggas were quite well striped on the body. Walter Rothschild obtained the specimen from Amsterdam in 1889. Despite visiting Tring earlier this year (and taking numerous photos), I failed to get a photo myself: this one is (c) Trustees of the Natural History Museum, London. The foal is a zebra x horse hybrid.
There are several ideas on how we should interpret this variation. The idea that the Quagga – when accepted as a species in its own right – might warrant splitting into several subspecies was once suggested but was abandoned by the 1970s (Groves 1974). More popular is the idea that this variation is due to Quaggas being part of a cline that’s continuous with variation seen in plains zebra populations to the north, this cline then being present within the Quagga itself; with this in mind, it makes sense that the least striped Quagga are the ones from the most southern limits of the animal’s range, while the stripiest are the ones closest to the range of Burchell’s zebra E. q. burchelli, the non-Quagga plains zebra population with the brownest, least stripey body. Incidentally, if you think of Burchell’s zebra as another extinct form, hold on: I have news which we’ll get to in a minute.
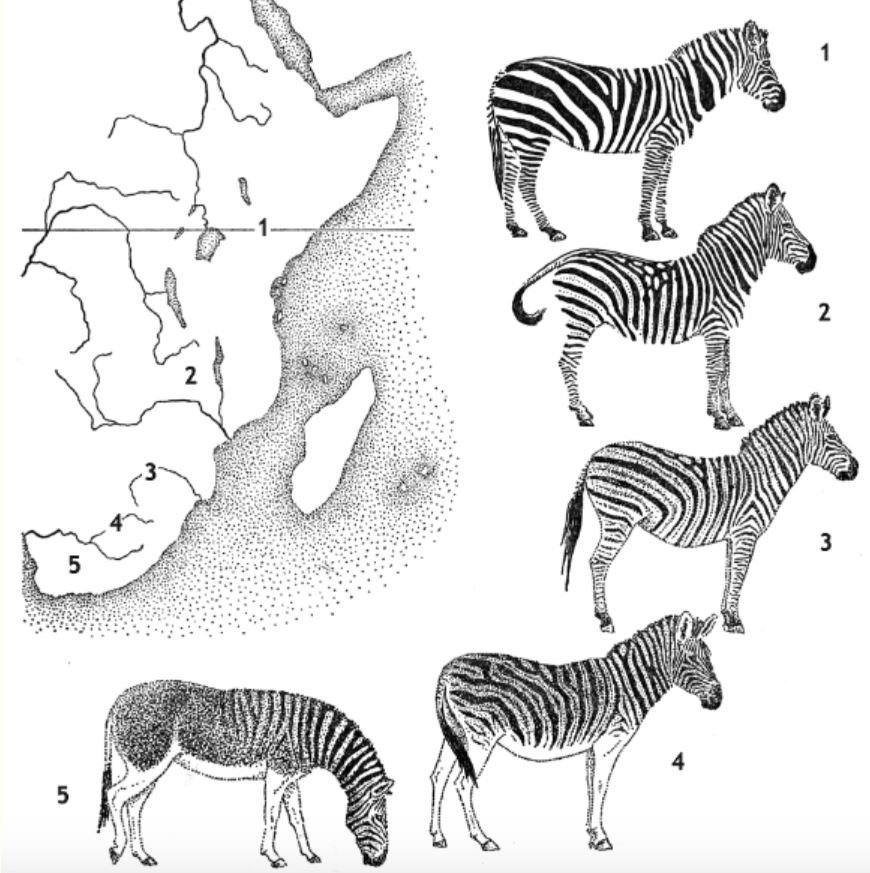
Caption: many of us are familiar with diagrams like this, which imply or outright aim to show that plains zebra form a cline of ‘decreasing stripiness’ from north to south. This image shows (1) Grant’s zebra, (2) Selous’s zebra, (3) Chapman’s zebra, (4) Burchell’s zebra and (5) Quagga; the map depicts where they occur. This image originally appeared in Andreas Suchantke’s 2001 book Eco-Geography; this version was modified by Craig Holdrege and is from this article at The Nature Institute.
In order to test this ‘clinal Quagga’ idea you’d need to analyse the distribution of the known Quagga specimens. But that can’t really be done (at least, based on present evidence) as locality data on their origins isn’t sufficiently precise. Those Quagga with the least amount of striping (historically named E. q. danielli) are known from far-flung locations across the Quagga’s range, and those with the most striping don’t definitely come from the north-east, as they ‘should’ according to the clinal hypothesis. A supposedly especially pale Quagga (conventionally E. q. isabellinus) is suggested to have lived in the area of the Lower Orange River in the west of the Quagga’s range, since mammals here tend to be pale (Groves 1974). This might be consistent with the cline hypothesis.
When it comes to how Quagga fit within plains zebra diversity as a whole, this cline idea is likely correct, but the irregular pattern of striping in the Tring Quagga and the variation in Quagga overall makes me wonder if Quagga were simply highly variable intraspecifically. Some genetic work indicates that Quagga evolved very recently – within the last 300,000 years (Leonard et al. 2005) – in which case their phenotype might not have been ‘fixed’. And if Quagga do/did form a cline with other plains zebra populations… well, that’s a little worrisome when it comes to the taxonomic distinction of these forms, since it implies that a fair degree of gene flow was happening in the zones of overlap. Hold that thought.

Caption: captive plains zebra at Marwell Wildlife, UK. If you know nothing about where they came from, how might these zebras be identified? Prominent, tall, striped manes, shadow stripes, reduced striping on the legs (especially in the animal on the right) suggest that they could be Burchell’s zebra, plus the body stripes aren’t as thick as those of Chapman’s zebra. But the five stripes that reach the belly are supposed features of Crawshay’s zebra. Image: Darren Naish.
Pale stripes, thin stripes, shadow stripes and more: variation in other plains zebras. Moving away from Quagga let’s move to the reasonable number of plains zebra populations (traditionally regarded as subspecies) that are most usually distinguished via the form, colour and spacing of their stripes. The taxonomic history of plains zebra is phenomenally complicated and numerous forms – initially proposed as species or subspecies – were named in the late 1800s and early 1900s. It didn’t take long for people to argue that many of the supposed distinguishing features of these alleged taxa were subject to substantial variation within populations (Cabrera 1936).

Caption: captive plains zebra at Marwell Wildlife, UK. As in the image above, I’m not sure which subspecies this animal is supposed to belong to. Note the ‘saddle’ formed by the transverse striping across the croup. Image: Darren Naish.
After an initial phase of confusion and the naming of umpteen forms, scientists eventually settled on the idea that plains zebra variation is consistent with the recognition of around five forms, all of which are best regarded as subspecies within Equus quagga (Groves & Bell 2003, Groves & Grubb 2011). With the Quagga regarded as an extreme ‘end member’ of a trend in stripe reduction, the general feeling in most discussions about plains zebra variation has been that the ‘ancestral’ plains zebras are those of the north-east, and that plains zebras become increasingly modified as we move south across the continent. Spoiler: this is likely completely wrong, but let’s hold fire yet.

Caption: bold and thick stripes with clean interspaces and relatively small size are characteristic of zebras identified as Grant’s zebra. This individual was photographed in Hell’s Gate National Park, Kenya. Image: Joachim Huber, CC BY-SA 2.0 (original here).
In the north-eastern part of the plains zebra’s range, we find the relatively small Grant’s zebra E. q. boehmi (böhmi in older literature), with thick, prominent and wholly black stripes, cleanly separated by white interspaces. A complex arrangement of stripes in the middle of the back is present in some individuals: a diagonal stripe heading from the croup (rump) toward the elbow is connected to a series of transverse stripes arranged across the back and croup. As a consequence, a sort of ‘saddle’ is formed. The body stripes in these zebras can be surprisingly thick. This is the archetypal zebra that most of us associate with the word zebra thanks to books, films and toys.
Occurring in Mozambique and nearby countries is the relatively obscure Selous’s zebra E. q. selousi, which is sometimes considered synonymous with E. q. borensis (selousi, also spelt selousii, was published in 1897 and thus wins out over borensis, named in 1921… if the two are regarded as synonymous). Groves & Bell (2004) recognised E. q. borensis as a taxon (Groves & Grubb (2011) called it the Half-maned zebra… read on) and didn’t recognise E. q. selousi. Anyway, these zebras have thinner stripes than Grant’s zebra and subtle shadow stripes across the body and hindlimb, and are sometimes characterised as having especially closely spaced, fine leg stripes all the way to the hooves. A ‘saddle’ of the sort described above for Grant’s zebra is sometimes present and is sometimes separated from the vertical stripes that connect the dorsal midline to the belly.
Occurring slightly to the west (in Zambia, Tanzania and Mozambique) is the also obscure Crawshay’s zebra E. q. crawshayi, which is also densely striped and with narrow interspaces. A supposedly unusual feature of Crawshay’s zebra is that five or more of its body stripes reach the belly (Groves & Bell 2004, Groves & Grubb 2011); supposedly, less than five stripes do so in other plains zebras. I’m very confused about this feature as I see it all over the place in plains zebras that aren’t meant to be Crawshay’s zebras. What am I misunderstanding?

Caption: animals identified as Crawshay’s zebra, at South Luangwa National Park (left) and an unspecified location (right). Note that relatively high number of stripes wrapping around the belly, and the feint shadow stripes present in the zebra on the right. Images: Prabir K. Bhattacharyya, CC BY-SA 3.0 (original here); Stfg, CC BY-SA 3.0 (original here).
Manelessness. In contrast to more familiar plains zebras (like Grant’s zebras), those zebras recognised as E. q. borensis – the Half-maned zebra of Groves & Grubb (2011) – don’t have a tall, prominent mane with black and white striping. The mane, in contrast, is either short and entirely black, or essentially absent. In some maneless zebras the ears are entirely white, and manelessness is clearly an evolutionarily derived condition as they start life with a mane but lose it as they mature. In some of these animals (like those of Sudan and Uganda), only males become maneless.
Caption: maneless zebra from Einar Lönnberg’s 1921 description of this form. The striping is crisp, prominent and complete but it looks like there are fine shadow stripes on the hindlimb.
Lest I create the impression here that reduced or absent manes are unique to Half-maned zebras, however, the actual situation is complicated: mane reduction and loss is present throughout east African plains zebra populations, occurring in those as far north as Somalia where the maneless zebras are supposed to belong to Grant’s zebra. There are definite maneless ‘centres’ (as in, areas where it’s especially prominent), but it seems to be a scattered phenomenon and not one that can be pinned on a specific taxon. What does this mean? Well, hang on.

Caption: maneless… or half-maned… zebra photographed at Liberec Zoo, Czech Republic. Maneless or half-maned zebras are in several zoos; in the UK, I’ve seen them at Colchester Zoo, Essex. Image: Jiří Sedláček - Frettie, CC BY-SA 3.0 (original here).
Chapman’s, Burchell’s and Damara or Damaraland zebras. Further south (from Angola to Namibia, Botswana, Zimbabwe and South Africa), there exists Chapman’s zebra E. q. chapmani, with paler, thinner black stripes and thin shadow stripes existing in the interspaces. It is said to have a proportionally large skull relative to other populations, and to have an especially short, narrow snout (Groves & Bell 2004, Groves & Grubb 2011).

Caption: Angel Cabrera’s 1936 illustrations of plains zebra variation. These are mostly variants of Burchell’s zebra but the five zebras at far left are Grant’s zebra and some at lower middle are Selous’s zebra. All are labelled individually in Cabrera’s paper, I’ve just thrown them together here. Image: Cabrera (1936).
And further south still (occurring from southern Botswana to Namibia in the west to far eastern South Africa in the east), there are – or were – plains zebras with mostly unstriped legs, thinner and paler black stripes and shadow stripes again. The interspaces in these animals are not as white as they are in plains zebras from further north; instead, they’re off-white or even yellowish. This is the aforementioned Burchell’s zebra.

Caption: the idea that Burchell’s zebra is extinct explains its appearance in various books that cover extinct mammals. This illustration by Maurice Wilson is from David Day’s 1981 The Encyclopedia of Vanished Species. It shows the reduced striping, prominent shadow stripes and mostly stripeless legs considered typical for this form. Image: Maurice Wilson.
Until recently it was thought that Burchell’s zebra was extinct, the last wild herds being killed off by 1910 and the last captive specimen dying in Berlin Zoo in 1918. However, Groves & Bell (2004) found that another southerly plains zebra ‘subspecies’ – the Damara or Damaraland zebra E. q. antiquorum – is continuous with the Burchell’s zebra morphologically, and hence that both should be united as a single taxon, the oldest name for which is E. q. burchelli Gray, 1824; it takes precedence over E. q. antiquorum (Smith, 1841). As emphasised by Groves & Grubb (2011), the type specimens of both burchelli and antiquorum came from adjacent regions in South Africa. These weren’t ever geographically or morphologically distinct taxa.

Caption: Burchell’s zebra in Etosha National Park, Namibia. Note the prominent shadow stripes, off-white interspaces, reduced leg striping, and presence of only three stripes contacting the belly. Image: Yathin S Krishnappa, CC BY-SA 3.0 (original here).
Striping: why so variable? Why might this variation in the size, intensity and distribution of stripes occur? Why zebras have stripes in the first place is – famously – one of the most-asked questions in zoology, and one with the largest number of proposed answers. If striping is (as has been suggested), connected to climate and thermoregulation, or the presence of biting flies or lions, it follows that variation in those things would lead to variation in striping.
My impression from the literature is that the thermoregulation hypothesis is currently winning out (Larison et al. 2015), rightly or not: the zebras with the boldest, darkest stripes are those from the hottest part of the plains zebra’s range while those with the most reduced striping are from the most arid, coolest parts (Leonard et al. 2005). But if stripes are this ‘adaptive’ relative to climate, what does this tell us about plains zebra variation? Could zebra populations that move into similarly warm – or cool – areas convergently develop similar striping? And… if so, does this negate the view (at least partially, anyway) that striping is taxonomically diagnostic? If there’s a cline – as implied above – isn’t the variation meant to be continuous? Remember also that manelessness – a feature implied here and there to be diagnostic for certain of the subspecies – occurs here and there across eastern plains zebra populations.
Caption: Quagga Project zebras at National Park, South Africa. These are Burchell’s zebras, with some being selectively bred for reduced striping. Image: Bernard Dupont, CC BY-SA 2.0 (original here).
The bombshell on this issue is a study that found plains zebras to group into nine populations that don’t conform to current taxonomy, and which have diversity maintained by extensive gene flow (Pedersen et al. 2018). This study found that plains zebras originated in southern Africa (probably in or around Botswana and Zambia), and from here spread north, north-east, and also south-east. The most divergent plains zebras – more divergent than even the Quagga – are the living plains zebras of Uganda (Pedersen et al. 2018).
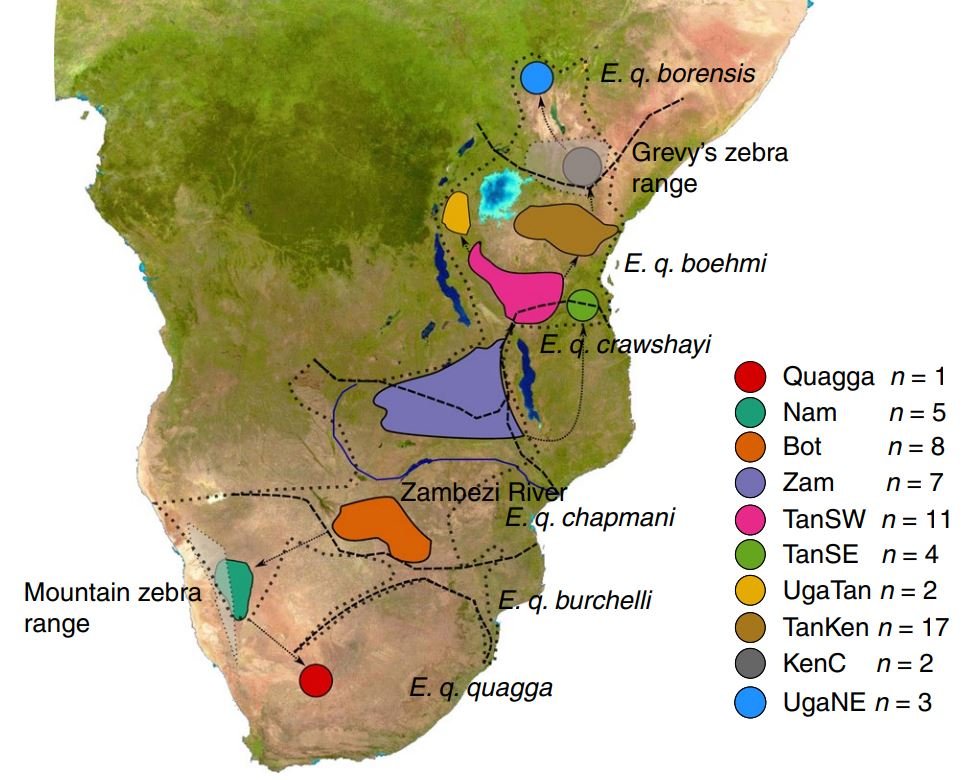
Caption: plains zebra range map from Pedersen et al. (2018). The dotted lines indicate the limits of plains zebra distribution, and the dashed lines show the boundaries between currently recognised subspecies. Note that the populations recognised in this study either overstep those boundaries, or occur within them.
South to north (not north to south) and the quest for a new taxonomy. What does this mean for our ideas about clinal variation within plains zebras, and what about taxonomy? Because Quaggas are – in this model – a southern extension of populations that occur further north (in Namibia and nearby), it’s still fair to regard Burchell’s zebra and the Quagga as part of a cline involving stripe reduction. But do the other plains zebra populations form a cline? Err, I’ll get back to you on that.
Rather than slim-striped, shadow-striped zebras like Chapman’s zebra being ‘advanced’ relative to densely-striped, thick-striped animals like Grant’s zebra, it might be the other way round. Maybe the zebras of the tropical north, with their super bold striping that extends all the way to the hooves and their occasional manelessness are the evolutionarily youngest and most anatomically modified.
We’re left with a situation whereby the anatomical (mostly stripe-based) variation that exists within the plains zebra may not relate to genetically distinct units at all, and that the genetically distinct units that do exist don’t conform to subspecies as currently understood. Pedersen et al. (2018) note specifically, for example, that populations previously labelled E. q. boehmi and E. q. crawshayi don’t group together, their conclusion being that these subspecies are artificial and polyphyletic. Geneticists sometimes imply from such results that we all need to throw up our arms, give up on existing taxonomic schemes, and devise something new. That’s not what we should do at all given that the holotypes of the innumerable named plains zebra taxa do belong to existing populations: in other words, even if you identify or ‘discover’ a new population, there’s almost certainly an appropriate name for it already. Getting to the bottom of that mess is not a fun or easy job though.

Caption: George Stubbs’s 1763 painting Zebra. It depicts a mountain zebra… on which more later.
And that’s where we’ll end for now. My original plan here was to write a single article that covers all the things I want to say about zebra variation. But it’s proved too much work, and consequently I’ve had to split things into several articles. More zebras soon.
For previous Tet Zoo articles on horses, wild and domestic, see…
A day at London’s Grant Museum of Zoology, August 2011 (includes discussion of Quagga and other zebras)
Fantastic asses, September 2013
Spots, Stripes and Spreading Hooves in the Horses of the Ice Age, February 2015
Domestic Horses of Africa, July 2022
Refs - -
Barnaby, D. 1996. Quaggas and Other Zebras. Basset Publications, Plymouth.
Bennett, D. K. 1980. Stripes do not a zebra make, part I: a cladistic analysis of Equus. Systematic Zoology 29, 272-287.
Cabrera, A. 1936. Subspecific and individual variation in the Burchell zebras. Journal of Mammalogy 17, 89-112.
Caro, T., Izzo, A., Reiner, R. C., Walker, H. & Stankowich, T. 2014. The function of zebra stripes. Nature Communications 5, 3535.
Crees, J. J. & Turvey, S. T. 2014. Holocene extinction dynamics of Equus hydruntinus, a late-surviving European megafaunal mammal. Quaternary Science Reviews 91, 16-29.
George, M. & Ryder, O. A. 1986. Mitochondrial DNA evolution in the genus Equus. Molecular Biology and Evolution 3, 353-546.
Groves, C. P. 1974. Horses, Asses and Zebras in the Wild. David & Charles, Newton Abbot and London.
Groves, C. P. & Bell, H. B. 2004. New investigations on the taxonomy of the zebras genus Equus, subgenus Hippotigris. Mammalian Biology 69, 182-196.
Groves, C. P. & Grubb, P. 2011. Ungulate Taxonomy. The Johns Hopkins University Press, Baltimore.
Higuchi, R., Bowman, B., Freiberger, M., Ryder, O. A. & Wilson, A. C. 1984. DNA sequences from the quagga, an extinct member of the horse family. Nature 312, 282-284.
Higuchi, R. G., Wrischnik, L. A., Oakes, E., George, M., Tong, B. & Wilson, A. C. 1987. Mitochondrial DNA of the extinct quagga: relatedness and extent of postmortem change. Journal of Molecular Evolution 25, 283-287.
Hvass, H. 1961. Mammals of the World. Methuen & Co, London.
Larison, B., Harrigan, R. J., Thomassen, H. A., Rubenstein, D. I., Chan-Golston, A. M., Li, E. & Smith, T. B. 2015. How the zebra got its stripes: a problem with too many solutions. Royal Society Open Science 2: 140452.
Leonard, J. A., Rohland, N., Glaberman, S., Fleischer, R. C., Caccone, A. & Hofreiter, M. 2005. A rapid loss of stripes: the evolutionary history of the extinct quagga. Biology Letters 1, 291-295.
Orlando, L., Metcalf, J. L. Alberdi, M. T., Telles-Antunes, M., Bonjean, D., Otte, M., Martin, F., Eisenmann, V., Mashkour, M., Morello, F., Prado, J. L., Salas-Gismondi, R., Shockey, B. J., Wrinn, P. J., Vasil’ev, S. K., Ovodov, N. D., Cherry, M. I., Hopwood, B., Male, D., Austin, J. J., Hänni, C. & Cooper, A. 2009. Revising the recent evolutionary history of equids using ancient DNA. Proceedings of the National Academy of Sciences 106, 21754-21759.
Pedersen, C.-E. T., Albrechtsen, A., Etter, P. D., Johnson, E. A., Orlando, L., Chikhi, L., Sigismund, H. R. & Heller, R. 2018. A southern African origin and cryptic structure in the highly mobile plains zebra. Nature Ecology & Evolution 2, 491-498.
Ray, R. E. 1978. Additions to the revised list of preserved material of the extinct Cape colony quagga and notes on the relationship and distribution of southern plains zebras. Annals of the South African Museum 77, 27-45.
Steiner, C. C., Mitelberg, A., Tursi, R. & Ryder, O. A. 2012. Molecular phylogeny of extant equids and effects of ancestral polymorphism in resolving species-level phylogenies. Molecular Phylogenetics and Evolution 65, 573-581.
Steiner, C. C. & Ryder, O. A. 2011. Molecular phylogeny and evolution of the Perissodactyla. Zoological Journal of the Linnean Society 163, 1289-1303.
Vilstrup, J. T., Seguin-Orlando, A., Stiller, M., Ginolhac, A., Raghavan, M., Nielsen, S. C., Weinstock, J., Froese, D., Vasiliev, S. K., Ovodov, N. D., Clary, J., Helgen, K. M., Fleischer, R. C., Cooper, A., Shapiro, B. & Orlando, L. 2013. Mitochondrial phylogenomics of modern and ancient equids. PLoS One 8 (2): e55950.



