Time to engage in more lark-themed musings of a phylogenetic nature….
Caption: lark images from the previous lark article. Images (clockwise from left): H. E. Dresser, in public domain; Darren Naish; J. G. Keulemans, in public domain.
You’ll no doubt have read the recent Tetrapod Zoology article on larks – A History of Larks: Twists and Turns and Overlooked Convergence – and thus assigned its various arguments and contentions to memory. Here, I want to circle back to what I said in that article about certain lark ‘species’ being species complexes. Buckle in, it’s quite a ride.
If you’re at all familiar with the diversity and taxonomy of living birds, you’ll know that a great many species are polytypic, consisting of some or many ‘subspecies’ that are often about as different from the nominal ‘subspecies’ as are supposedly distinct, related species within the relevant group. The subspecies issue is often treated as a sort of joke, as if subspecies have been invented by bureaucrats for book-keeping purposes or by birdwatchers desperate to increase their list of ticks in a notebook.

Caption: you might get the impression from all the complaining that subspecies are some irksome ever-present annoyance that you can’t escape from. They’re really not. Books like these only ever include data on a handful of them, and if you actually want to know what’s going on you have to dig deep into the technical literature. Image: Darren Naish.
‘Subspecies’ 1: mostly a waste of time. On the negative side of things, it is indeed the case that quite a few ‘subspecies’ don’t prove worthy of recognition when we study the relevant populations in detail. Those supposed subspecies that can be differentiated due to slightly larger or smaller size, or lighter or darker plumage, sometimes seem instead to be end members of a cline, and they prove unsupported as distinct entities when their genetics are examined. I’m not old enough to really remember a time when British birds were treated as endemic ‘subspecies’, but I do own books from the 1950s and 60s where at least some British populations of Eurasian species are treated as such. Today, we mostly laugh at that idea (Hebridean wrens excepted).

Caption: Charles Tunnicliffe’s 1965 Wild Birds of Britain is a wonderful piece of work. But it was written at a time when people were still thinking that even the most minor of anatomical differences were sufficient for a population to be differentiated as a ‘subspecies’. Are British Goldcrests Regulus regulus really distinct enough from those of mainland Europe to be worthy of subspecific recognition? Today, we say no.
Examples in larks of defunct subspecies include the many supposed local forms of Greater short-toed lark Calandrella brachydactyla and Red-capped lark C. cinerea, most of which have not been found to be supported by cytochrome b analysis (Stervander et al. 2016).
‘Subspecies’ 2: actually good, worthy, and deserving of recognition. Having said all that, the fact remains that some and perhaps many subspecies are actually ‘distinct’. If you adhere to a ‘diagnostic species concept’ that’s combined with studies of phylogeny, those entities we term ‘species’ are those biological units that can (1) be defined and differentiated on the basis of diagnostic traits and (2) form ‘distinct lineages’ in the phylogeny, which is to say that they’re not nested within another unit already recognised as a species and diverged at an estimated point in geological history consistent with species status. Take this as far as it can go, and many ‘subspecies’ should actually be regarded as ‘species’: a decision that might require us to approximately double the recognised number of living bird species (Zink 1996, 2004, Zink & McKitrick 1995, Barrowclough et al. 2016).
For all those claims that increasing the number of ‘species’ is due merely to ‘taxonomic inflation’ and changing species concepts, the counter-argument is that the relevant decisions have been based on appropriate study and newly acquired data (Sangster 2009). That’s obvious if you go to the trouble of digging into the technical literature. Modern papers that make taxonomic recommendations about species boundaries involve a substantial amount of hard work.
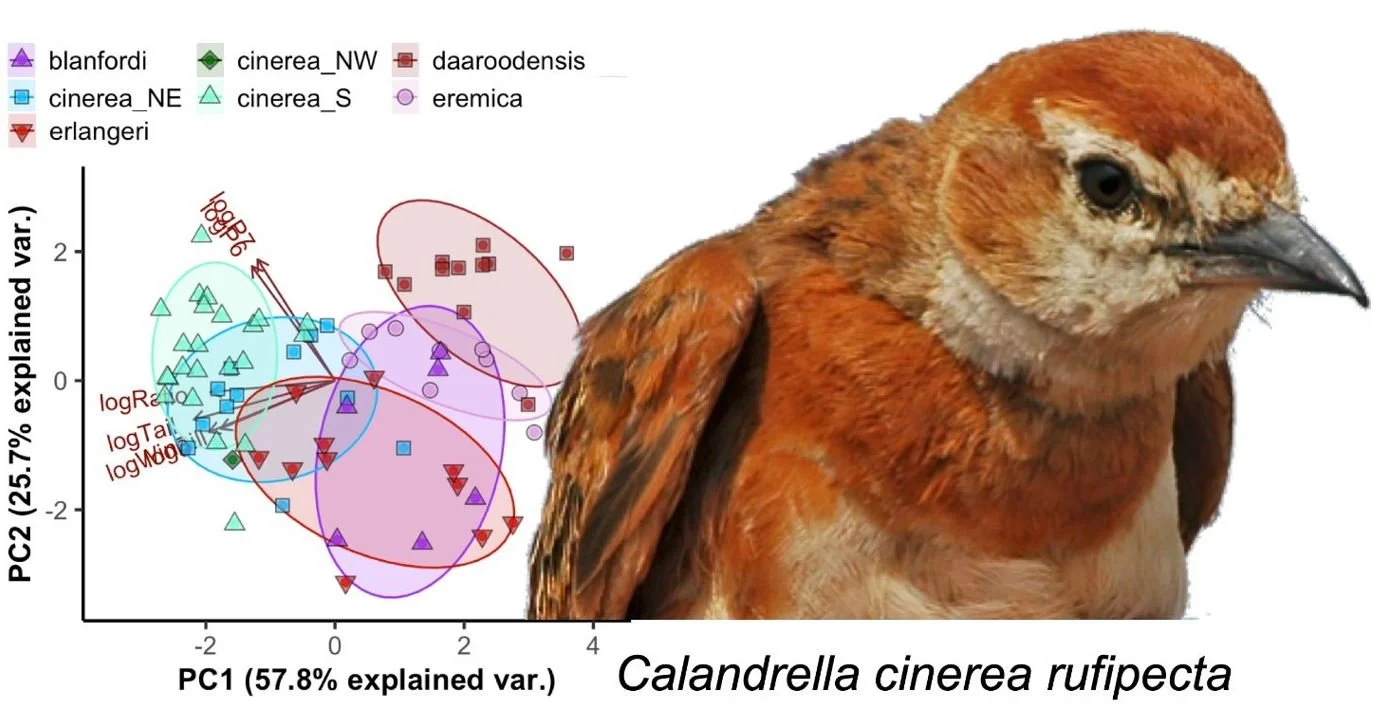
Caption: mostly gone are the days where you can just dismiss subspecies as local variants invented by people who never leave museums. Modern analyses of subspecies-level variation involve morphometrics, DNA and data on range and ecology. The new Red-capped lark subspecies C. cinerea rufipecta was named by Stervander et al. (2020).
Larks: cryptic species and rampant paraphyly. Let’s take this back to larks again. Recent studies have shown that at least some taxa once considered ‘subspecies’ warrant recognition as species, both because they’re diagnostic, and because they’ve been found to be ‘distinct lineages’ as explained above.
Stervander et al. (2016, 2020) looked at populations within Calandrella larks, and found Greater short-toed lark C. brachydactyla, Red-capped lark C. cinerea, Hume’s short-toed lark C. acutirostris and others to have “unexpected deep divergences” (meaning that they originated a surprisingly long time ago within the geological history of the group), and to sometimes be paraphyletic with respect to other Calandrella species. The data revealing “unexpected deep divergences” requires that certain of the lineages conventionally included within the species concerned diverged during the Zanclean Age of the Pliocene (about 4 million years ago), or even before this, meaning that they almost certainly represent cryptic species.
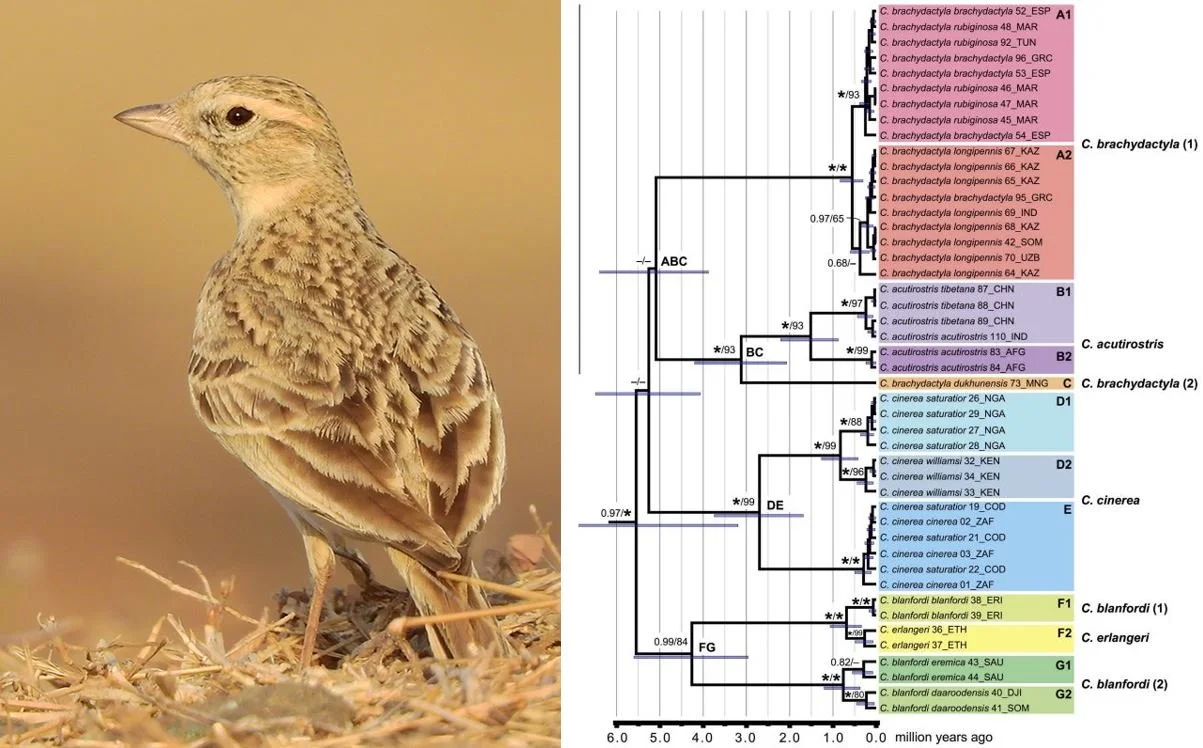
Caption: Greater short-toed lark photographed in Maharashtra, India. At right, Calandrella lark phylogeny from Stervander et al. (2016), based on mitochondrial gene b. Of note here is that some subspecies (like C. brachydactyla dukhunensis, shown in orange) don’t group with the rest of their species, and that some subspecies-level divergences (like that between C. b. dukunensis and the rest of C. brachydactyla, and those within C. blanfordi) are posited to have occurred during the Pliocene. Images: Shantanu Kuveskar, CC BY-SA 4.0 (original here); Stervander et al. (2016).
An east Asian lark conventionally included within the Greater short-toed lark has been shown in several studies to be closer to Hume’s short-toed lark and be very distant on the tree from the Greater short-toed lark. It’s now generally regarded as a distinct species: the Mongolian short-toed lark C. dukhunensis (Alström et al. 2013, 2020).
Ghorbani et al. (2020) found populations conventionally grouped together within the Lesser short-toed lark Alaudala rufescens* to form four clades that diverged from one another between about 1.6 and 3.2 million years ago, their conclusion being that there are probably four cryptic species here. Furthermore, the Sand lark Al. raytal is nested within this complex, meaning that the traditional version of the Lesser short-toed lark is paraphyletic (Ghorbani et al. 2020).
* Alaudala rufescens was conventionally termed the Lesser short-toed lark. But since the publication of the studies discussed here, it’s become increasingly popular to refer to A. rufescens in the new, restricted sense as the Mediterranean short-toed lark.

Caption: you don’t need to see the whole cladogram (its top is chopped off) to get the point here. Ghorbani et al. (2020) found the animals conventionally included together within Alaudala rufescens to consist of four clades, with the Sand lark Al. raytal nested in among them. Once again, note that the divergence times of these clades occurred at times ordinarily deemed ‘about right’ for species-level divergences. Image: Ghorbani et al. (2020).
Finally, Stervander et al. (2020) found deep evolutionary divergences within species of Eremophila (horned larks), Galerida (crested larks), Alauda (skylarks), Alaemon (hoopoe-larks), Eremopterix (sparrow-larks), and in others. Species in all of these groups include ‘subspecies’ that diverged more than 2, or more than 3, or even more than 5 million years ago. These divergences corresponded to biogeographical barriers, are “mirrored by several concordantly distributed mammalian savanna specialists” (Stervander et al. 2020, p. 17), and sometimes concern isolated populations that appear to be relicts* of occurrence across a broader, ancestral habitat.
* Stervander et al. (2020) used the word ‘relic’ but that doesn’t seem right to me. Isn’t ‘relic’ more associated with the leftovers of human civilisations? I’m more used to seeing organisms being referred to as ‘relictual’.

Caption: at left, a Mediterranean short-toed lark Alaudala rufescens photographed in Israel. At right, a Sand lark Al. raytal photographed in India. Genetics indicates that Al. raytal is nested within the traditional version of Al. rufescens where it’s one of five deep, distinct lineages. The solution most consistent with decisions made elsewhere in passerine taxonomy is that all five should be retained as species or elevated to species level. Larks of many sorts can raise and lower the feathers on the crown as desired, and thus end up with different profiles one moment to the next. Images: Lior Kislev, CC BY-SA 4.0 (original here); Savithri Singh, CC BY-SA 4.0 (original here).
Take-home. I know this can be hard to follow if you don’t know all the species/subspecies in question, but the take-home messages here are pretty simple. And while I’ve concentrated here on larks, they’re echoed in numerous other studies of passerine groups. Despite what you’ve heard and read about zoologists – and ornithologists in particular – being trigger-happy splitters, overly keen to recognise ‘species’ when a simpler, lazier, lumpier approach will suffice, the fact is that many (perhaps most) polytypic ‘species’ aren’t monophyletic when studied in detail. The more distinct of their supposed ‘subspecies’ don’t belong with the nominal form and go elsewhere in the tree.
Reticulation and hybridisation introduce further complexity to this issue, for sure, but are not immediately relevant to the cases discussed in this article.
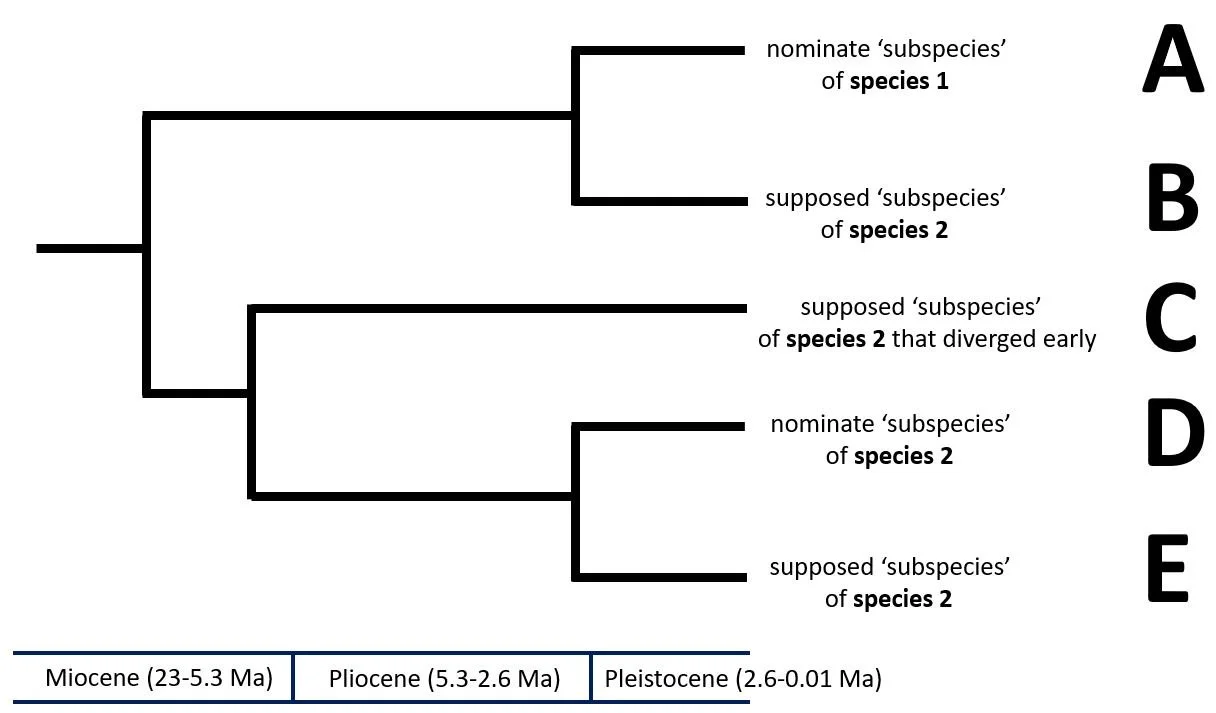
Caption: here’s a hypothetical time-calibrated cladogram that illustrates the sorts of issues we’re dealing with. How do we treat (in the taxonomic sense) taxa B, C and E? The primary assumption here is that taxa that have been separate for more than c 2 Ma are ‘different enough’ to be species. B seems here not to belong to the same species as D (as had previously been thought), so is it a new species, or is it part of the same species as A? Is C a distinct species, or is it part of the same species as D and E? And is even E ‘distinct enough’ from D to be a distinct species? After all, the two diverged in the Pliocene.
It’s not just ‘species’ that are non-monophyletic. A vast number of studies now (referring to passerine birds here) show that traditional genera are paraphyletic or monophyletic, and are little more than ‘form taxa’ when analysed rigorously. Do we adapt to this by adopting a very ‘lumpy’ taxonomy, or do we do the opposite and resurrect many old names and coin many new ones? Here, the former approach is mostly winning out.
In which we end by beating up Carl Linnaeus. I’ll end by saying that all of this constant chopping and changing to taxonomy requires that writers and researchers have to check which species- or genus-level name is the ‘correct’ one at the time of writing. And a lot of people don’t like this, the main complaint being that they’d prefer if it taxonomic labels just stayed the same.
I have two points in response to that. Point 1 is that the Linnaean binomial system is a bad one, and that we’ve done ourselves no favours by sticking with it. Linking taxonomy (the naming of things) with phylogeny (the determining of the relatedness of things) was always a bad idea, since our phylogenetic models are constantly in flux. They always have been, and they always will be. We’re thus stuck with a system where taxa will always be moving around, and where names will need to be changed.

Caption: oh Carl, what have you done? Image: Nationalmuseum (Stockholm), public domain (original here).
Point 2 is that this changing is a consequence of people learning new stuff and doing new studies. So long as we keep learning new stuff about phylogeny and thus about the phylogenetic position of organisms, we’ll need to keep modifying the taxonomy. If you’re interested in nature and science, you presumably want people to keep doing scientific research. Ergo, complaining about the labile nature of taxonomy (see Point 1) can come across as anti-scientific (see Point 2).
Is there an alternative? Yes. Devise a new system where taxonomy is not connected to phylogeny. Could such a system replace the existing one, and is it really worth the hassle at this point? Dear reader, I leave you to decide.
For previous Tet Zoo articles on passerines, see…
Great tits: still murderous, rapacious, flesh-rending predators!, February 2013
For The Love of Crows, October 2015
Thoughts on the Passerine Tree, 2016, October 2016
A Battle Among Blue Tits, February 2018
Cocks-of-the-Rock, Extreme Cotingas, April 2019
Birdwatching in Suburban China, May 2019
Tell Me Something Interesting About Dunnocks, July 2019
Suburban Birdwatching in Queensland, Australia, November 2019
Kirk W. Johnson’s 2018 The Feather Thief, a Review, February 2021
The Locustella Warblers, May 2021
Chiffchaffs and What Are 'Old World Warblers' Anyway?, February 2022
Surprising Diet of the House Sparrow, August 2022
A History of Larks: Twists and Turns and Overlooked Convergence, February 2023
Refs - -
Alström, P., Barnes, K. N., Olsson, U. Barker, F. K., Bloomer, P., Khan, A. A., Qureshi, M. A., Guillaumet, A., Crochet, P. A. & Ryan, P. G. 2013. Multilocus phylogeny of the avian family Alaudidae (larks) reveals complex morphological evolution, non-monophyletic genera and hidden species diversity. Molecular Phylogenetics and Evolution 69, 1043-1056.
Alström, P. & Sundev, G. 2020. Mongolian short-toed lark Calandrella dukhunensis, an overlooked East Asian species. Journal of Ornithology 162, 165-177.
Barrowclough, G. F., Cracraft, J., Klicka, J. & Zink, R. M. 2016. How many kinds of birds are there and why does it matter? PLoS ONE 11, e0166307.
Ghorbani, F., Aliabadian, M., Zhang, R., Irestedt, M., Hao, Y., Sundev, G., Lei, F., Ma, M., Olsson, U. & Alström, P. 2020. Densely sampled phylogenetic analyses of the Lesser Short-toed lark (Alaudala rufescens) — Sand lark (A. raytal) species complex (Aves, Passeriformes) reveal cryptic diversity. Zoologica Scripta 49, 427-439.
Sangster, G. 2009. Increasing numbers of bird species result from taxonomic progress, not taxonomic inflation. Proceedings of the Royal Society of Biological Sciences, Series B 276, 3185-3191.
Stervander, M., Alström, P., Olsson, U., Ottosson, U., Hansson, B. & Bensch, S. 2016. Multiple instances of paraphyletic species and cryptic taxa revealed by mitochondrial and nuclear RAD data for Calandrella larks (Aves: Alaudidae). Molecular Phylogenetics and Evolution 102, 233-245.
Stervander, M., Hansson, B., Olsson, U., Hulme, M. F., Ottosson, U. & Alström, P. 2020. Molecular species delimitation of larks (Aves: Alaudidae), and integrative taxonomy of the genus Calandrella, with the description of a range-restricted African relic taxon. Diversity 12 (11), 428.
Zink, R. M. 1996. Bird species diversity. Nature 381, 566.
Zink, R. M. 2004. The role of subspecies in obscuring avian biological diversity and misleading conservation policy. Proceedings of the Royal Society of London B 271, 561-564.
Zink, R. M. & McKitrick, M. C. 1995. The debate over species concepts and its implications for ornithology. The Auk 112, 701-719.



