Dicynodonts are quite odd. But what if they were even… odder?

Caption: dicynodonts are great, and I totally haven’t written enough about them. This is Sinokannemeyeria from the Middle Triassic of China, on display at the Paleozoological Museum of China. Image: Morosaurus millenii, CC BY-SA 4.0 (original here).
I’ve definitely said on several previous occasions that stem-mammals – that huge assemblage of extinct tetrapods that used to be called ‘mammal-like reptiles’ – have never received due coverage here; I’ve just never had the time, nor been equipped with sufficient imagery, to write about them all that much. This will change in the future, but that’s a thing I’ve been saying for more than ten years now.
Anyway, I do want to talk about stem-mammals today, but only about one very specific, very niche aspect of their portrayal in the palaeontological literature. I refer to the idea (only ever mentioned a handful of times) that some big dicynodonts might have been capable of bipedal rearing and might have behaved something like certain ground sloths.

Caption: in which I try as hard as I can to make a kannemeyeriiform dicynodont - in this case, Dinodontosaurus from the Middle Triassic of Brazil - to look bipedal in sloth-like fashion. Even those animals absolutely unsuited for bipedal rearing can still do it on occasion, so maybe this scene isn’t thoroughly implausible. But were kannemeyeriiforms specialised for this behaviour? Image: Darren Naish.
Dicynodonts are quadrupedal, heavily-set, short-tailed synapsids best known for their beaked, (typically) tusked faces. They were a diverse bunch that got up to an awful lot of things during their long evolutionary history. There were relatively small, short-legged, burrowing dicynodonts, some archaic, toothed, mid-sized taxa that look like they were omnivorous generalists, and a wide variety of large and very large browsing taxa that often combined a deep, toothless beak with big, descending maxillary flanges. We used to say that the biggest of these animals (like Kannemeyeria) had a skull length of around 40 cm long, an overall length of around 3 metres and a weight of something like 400 kg. But that was before the 2018 publication of Lisowicia from the Late Triassic of Poland, a giant over 2 metres tall at the hips, 4.5 metres in length (or more), and a weight of over 4 tons.

Caption: a dicynodont montage produced for my in-prep textbook (which can you learn more about, and support, here). Image: Darren Naish.
There’s also a long-standing suggestion that some taxa (most famously the very widespread and speciose Permo-Triassic Lystrosaurus) might have been amphibious. While I don’t have anything in particular against that idea, I’ve never understood why it’s been so popular and suspect that it’s another of those cases where palaeontologists see an animal with a barrel-shaped body and assume it must have lived like a hippo. It’s a pretty crass approach to determining palaeoecology and often, I think, erroneous. Having said that, some experts have argued that bone microtexture does in fact demonstrate that Lystrosaurus was amphibious (Ray et al. 2005). But that’s a story for another time.
A far less familiar idea about dicynodont behaviour is that at least some species included within the large-bodied Triassic* group Kannemeyeriiformes might have reared up bipedally to feed from branches growing perhaps two or three metres off the ground. Say whaaat??
* Actually, kannemeyeriiforms surely first appeared in the Late Permian, but we currently lack fossils from this early part of their history.

Caption: my reasonably well-used copy of King (1990). Image: Darren Naish.
I first learnt of this idea from Gillian King’s 1990 book The Dicynodonts: A Study in Palaeobiology (which I obtained at bargain price – the book is infamously expensive – due to the kindness of a friend) and was shocked – shocked, I tell you – to see it put forward. King explained that it originated in a 1978 paper by the (now no longer with us) Arthur Cruickshank, which I was also able to obtain. The paper in question – ‘Feeding adaptations in Triassic dicynodonts’ (Cruickshank 1978) – begins by making some broad-brush comparisons between the overall shape of kannemeyeriiforms and the widths of their snouts with those of living herbivores and herbivore-omnivores, namely pigs and rhinos. This was very much the approach published by Cox (1965) who compared (relatively) narrow-snouted dicynodonts with browsers like the Black rhino Diceros bicornis and (relatively) broad-snouted dicynodonts with grazers like white rhinos Ceratotherium. There are a bunch of problems with those comparisons, they’re probably erroneous, and this area has been revisited in recent years by other researchers; I might discuss this area another time.
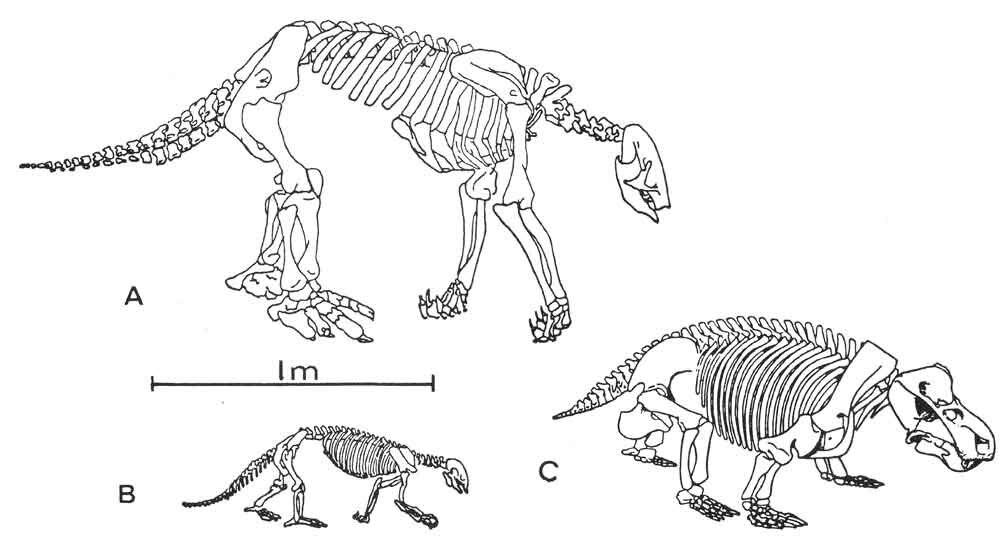
Caption: Cruickshank (1978) compared the skeletons of dicynodonts like (C) Kannemeyeria with sloths, like (A) Nothrotherium and (B) Hapalops. Image: Cruickshank (1978).
What makes Cruickshank (1978) novel is that he pointed to several unusual features of kannemeyeriiforms as indicative of an unusual feeding behaviour. He suggested that the reinforced skull roof “could be an adaptation for pushing over vegetation” (p. 124) and that the long claws on the forelimbs (what – really? I didn’t know kannemeyeriiforms had long claws), capable of extreme flexure, “could act as powerful hooks to pull leaf bunches to the mouth” (p. 126). From here, he noted that ground sloths might be analogues of the dicynodonts in question, and he went on to say that kannemeyeriiform and ground sloth postcranial skeletons were “strikingly convergent”.
In the accompanying diagrams, Cruickshank drew attention to the similarity of sloth and kannemeyeriiform hindlimbs, humeri, pelvic girdles, hands and lower jaws (Cruickshank 1978). A channel present along the floor of the mandibular symphysis in the African kannemeyeriiform Dolichuranus led him to suggest that these animals had a long, prehensile tongue – another feature seen as sloth-like. And in a life reconstruction included within the paper (penned by Dolores Rokos) – controversially depicting some Sinokannemeyeria-like animals with shaggy coats – kannemeyeriiforms were shown as knuckle-walkers (Cruickshank 1978). That’s a bold thing to show, since habitual knuckle-walking (which actually involves animals walking on their flexed digits, not on the structures I think of as knuckles) is a specialised behaviour which requires a specialised anatomy.

Caption: the rather curious depiction of living kannemeyeriiforms featured in Cruickshank (1978); illustration by Dolores Rokos. The shaggy fur is obvious, but look also for the prehensile tongues, unusual hand and foot posture, as well as the bipedal rearing. Image: Cruickshank (1978).
Interesting, and radical, stuff, mostly unreported in what little popular literature discusses dicynodonts and their biology. How did this idea fare in the years following its proposal? As you might guess from the fact that it appears to have sunk without a trace… it was mostly ignored, and I think this is because most other specialists realised (or knew) that it wasn’t really supported by the anatomical evidence and hence didn’t warrant response. Here we must take a brief diversion.

Caption: a close-up of one of those shaggy kannemeyeriiforms from Cruickshank (1978).
On heterodox ideas and what to do with them. As I’ve said here before, there are two schools of thought on what scientists should do when observing the publication of an unusual, heterodox, and likely incorrect, claim. School Number One is that said claim should be ignored and simply not mentioned or cited, since citing or discussing something brings attention to it (thereby boosting its visibility and citation metrics), and – if ignored – it will simply sink into the morass of obscurity and be forgotten forever. School Number Two is that said claim should be definitively smacked down with great vengeance and furious anger in order that it not poison the minds of lesser readers who may encounter the claim and think that it actually represents a good argument, or ‘the truth’.
The dicynodont literature shows that School Number One was followed, rightly or not. Indeed, just about the only published response available (until recently; read on) on Cruickshank’s proposal is included within a review of King’s 1990 book, namely that published by dicynodont expert Laurie Walter (Walter 1991). Walter liked King’s book, but one thing she didn’t like was that King (1990) was “simply insufficiently critical” of palaeoecological proposals made for the group, in particular those put forward by Cruickshank (1978). Walter dismissed Cruickshank’s sloth-like idea as “fanciful” and noted that one of the key morphological similarities which Cruickshank (1978) drew attention to (that concerning the pelvis) was bogus: Cruickshank was right that ground sloths and kannemeyeriiforms both have a flaring pelvis, but “[t]he expansions are … in different planes in the two groups: the ilia are splayed laterally in ground sloths, forming a plausible ‘basin’ for the digestive tract when the body is upright, but are longest anteroposteriorly in kannemeyeriids, ill-suited for visceral support in an upright, or any other, posture” (Walter 1991, pp. 261-262).

Caption: ground sloths as seen from behind (Eremotherium on the left, Glossotherium on the right). The funnel-shaped ribcages, massive flaring, thickened pelvic girdles, absurdly broad femora and other features are simply not like anything seen in kannemeyeriiforms or other dicynodonts. This photo was taken at the National Museum of Brazil, Rio de Janeiro, in 2013 (and thus prior to the fire of 2018). Image: Darren Naish.
While looking at ground sloth skeletons (as I have on a great many occasions, both at the several specimens in British museums as well as at those in the Brazilian and US collections I’ve visited), I’ve often had this alleged similarity with kannemeyeriiforms in mind, since – like Walter – it’s always struck me that sloths and kannemeyeriiforms are not similar at all, at least not in meaningful detail. It might be true that kannemeyeriiforms share with ground sloths robust limb bones and a broad pelvis, but these features have evolved multiple times among animals that differ markedly in ecology and lifestyle. Furthermore, the similarities between ground sloths and kannemeyeriiforms in these parts are totally superficial.

Caption: the fossil sloth that the majority of us are most familiar with, the giant South American megatheriid Megatherium americanum, an animal with a very important role in the history of vertebrate palaeontology. This specimen – a cast, on show at London’s Natural History Museum – has a fascinating backstory of its own, and has been on show in the museum since its opening in 1881. Image: Darren Naish/NHM.
The Cruickshank (1978) claim that kannemeyeriiforms have long hand claws which look suited for hooking onto foliage is also just not true. Look at the hand skeleton of Tetragonius which Cruickshank (1978) himself figured: the unguals are flattened and rounded, not hook-like or at all claw-like. The kannemeyeriiform hand is also built for typical plantigrade placement of the digits on the substrate of the sort seen in other dicynodonts (where it matches trackways): these animals were not ‘knuckle-walking’ as per the illustration in Cruickshank (1978).
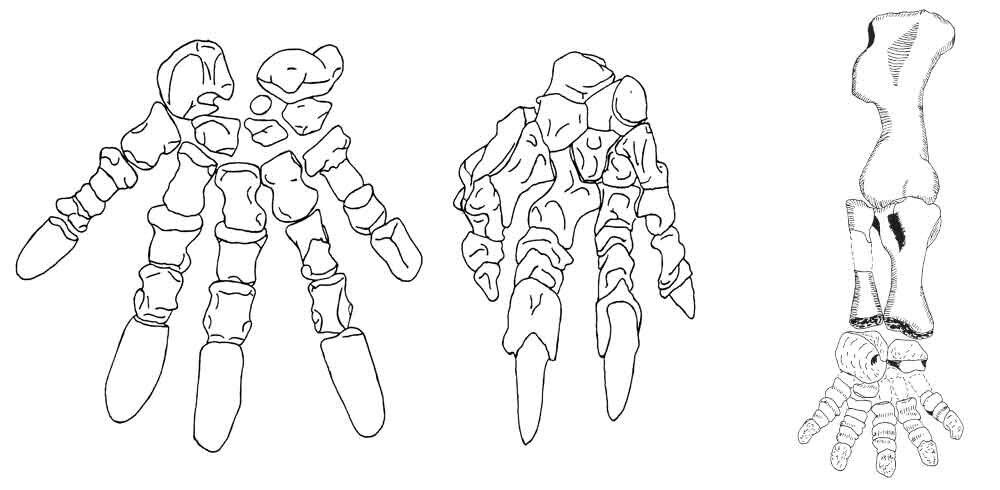
Caption: do kannemeyeriiform hands really look adapted for knuckle-walking, and do they have long, sloth-like claws? As you can see here, they don’t. At far left, we see the left hand of the kannemeyeriiform Tetragonius; in the middle is the right hand of the sloth Mylodon; and at far right is the right hindlimb of the kannemeyeriiform Ischigualastia. Images: Cruickshank (1978).
Furthermore, those ground sloths usually thought of as part-time bipeds (like the giant Megatherium) possess features which appear linked with bipedal habits (and we know that they sometimes moved bipedally, thanks to fossil tracks). These include a posteriorly located centre of mass, especially tall lumbar vertebrae, a freakishly broad, almost rectangular femur, proportionally long feet and a massive, muscular tail (Casinos 1996, McDonald 2007, Morato et al. 2008). Kannemeyeriiforms (and other dicynodonts) lack these features. In fact, the kannemeyeriiform skeleton looks utterly unsuited for bipedality of any sort. As noted right at the start of this article, this doesn’t mean that they couldn’t assume bipedal poses on occasion, but they sure weren’t specialised for bipedal standing or walking.

Caption: Morato et al. (2008) constructed a small model of the kannemeyeriiform Dinodontosaurus, scanned it digitally, and then used software to determine the centre of mass: it’s visible as the white dot. Image: Morato et al. (2008).
Thanks to synapsid expert Christian Kammerer, I was made aware of Morato et al. (2008), a paper which specifically tested Cruickshank’s ground sloth idea by looking at the position of the kannemeyeriiform centre of mass and the bending strength of their limb bones. Their conclusions? That the centre of mass was located in the thorax, close to the massively muscular, heavy forelimbs and pectoral girdle, and that their hindlimb bones were not exceptional in bending strength relative to those of the forelimb (in contrast to sloths like Megatherium) (Morato et al. 2008).

Caption: Morato et al. (2008) used a second technique to determine the position of the centre of mass in both juvenile and adult specimens of the kannemeyeriiform Dinodontosaurus: they constructed clay models and suspended them by wires. The results essentially match the digital technique illustrated above. Image: Morato et al. (2008).
Did, then, those big, kannemeyeriiform dicynodonts stand up bipedally in ground sloth fashion? No, they really did not. You might argue – as per School Number One option discussed earlier – that this was such a poor idea that it never really was worthy of extensive discussion. But I don’t quite agree. I think it was interesting enough, and novel enough, to get a fair hearing, and it resulted in some odd artwork – an aspect of the reconstruction process which I can’t help but pay attention to.
So is this the end of this ground sloth / kannemeyeriiform dicynodont cross-over thing? No, no it is not! We’ll return to this topic again in time…
For relevant previous articles on stem-mammals and other synapsids, see…
Survivors, diggers, herbivores, first giant terrestrial vertebrates: the caseids, July 2007
The answers we seek: on ‘goodbyes’, the necks of caseids, and weird mystery sauropods, July 2007
Perentie tries to swallow echidna. Echidna too spiky, Perentie gets horribly injured. Dies., December 2009
Late Cretaceous Animals of Romania's Haţeg Island--a More Complex View, September 2015
The Stem-Mammals--a Brief Primer, September 2016
Did Mesozoic Mammals Give Birth to Live Babies or Did They Lay Eggs?, May 2020
Refs - -
Casinos, A. 1996. Bipedalism and quadrupedalism in Megatherium: an attempt at biomechanical reconstruction. Lethaia 29, 87-96.
Cox, C. B. 1965. New Triassic dicynodonts from South Africa, their origins and relationships. Philosophical Transactions of the Royal Society of London B 248, 457-516.
Cruickshank, A. R. I. 1978. Feeding adaptations in Triassic dicynodonts. Palaeontologia Africana 21, 121-132.
King, G. 1990. The Dicynodonts: A Study in Palaeobiology. Chapman & Hall, London, New York.
McDonald, H. 2007. Biomechanical inferences of locomotion in ground sloths: integrating morphological and track data. New Mexico Museum of Natural History and Science Bulletin 42, 201-208.
Morato, L., Schultz, C.L., Vega-Dias, C., Pinto Da Silva, F. & Kindlein, W. 2008. Discussing a myth: biomechanical comparisons between Dinodontosaurus (Synapsida, Dicynodontia) and extinct ground sloths. Arquivos do Museu Nacional, Rio de Janeiro 66, 145-154.
Ray, S. 2006. Functional and evolutionary aspects of the postcranial anatomy of dicynodonts (Synapsida, Therapsida). Palaeontology 49, 1263-1286.
Ray, S., Chinsamy, A. & Bandyopadhyay, S. 2005. Lystrosaurus murrayi (Therapsida, Dicynodontia): bone histology, growth and lifestyle adaptations. Palaeontology 48, 1169-1185.
Walter, L. R. 1986. The limb posture of kannemeyeriid dicynodonts: functional and ecological considerations. In Padian, K. (ed) The Beginning of the Age of Dinosaurs. Cambridge University Press (Cambridge), pp. 89-97.
Walter, L. R. 1991. [Review of] The Dicynodonts: A Study in Paleobiology. Journal of Vertebrate Paleontology 11, 261-262.



