As a regular denizen of the TetZooniverse, you may well remember the July 2017 article ‘Fossils We Want to Find’ in which I discussed a list of hypothetical fossil things that we might one day discover but haven’t yet. Wouldn’t it be fun to do the same sort of thing with extant species; that is, with discoveries pertaining to living, breathing animals? Over at the Zoology for Enthusiasts facebook group (a spinoff of the Tetrapod Zoology facebook group), Jordan Fryer suggested doing exactly this, and consequently people have been coming up with their own suggested living animals that might await discovery. Because this seemed like a lot of fun (and a chance to discuss some really neat and unusual stuff), I thought I’d give it a go.

Caption: the 2017 precursor to the article you’re reading here was all about fossil animals. It included this photo, which shows me in the act of discovering a dinosaur bone in the Moroccan Sahara. Image: Richard Hing.
Naturally, any list of this sort is horribly subjective, reflecting the interests and biases of the person compiling the list, but so be it. It also seems all too easy to turn any compilation into a ‘list of most discoverable cryptids’: for those of you who don’t know, I have a long-standing interest in cryptozoology and have published on it quite frequently (see Naish (2017) for starters). For the most part, I’ve not done this, though read on.

Caption: many of my thoughts on mystery animals can be found in my 2017 book Hunting Monsters. I am not - sorry - much impressed by the case for such supposed animals as the mokele-mbembe, an artistic reconstruction of which is shown at right. Image: David Miller, in Mackal (1987).
I’ve also mostly excluded hypothetical discoveries that are inspired by the creatures of cryptozoology but could arguably be considered independent of the cryptozoological literature. In part this is because I don’t think they’re plausible or worth considering, but it’s also because they’re cliched and the opposite of original. So, no ‘living sauropods from the Congo’ or ‘living plesiosaurs in Loch Ness’, for example.
As for what I have selected: well, some of my suggestions are sillier than others, and some are perhaps not that interesting to non-specialists. But, whatever. Feel free to dissect my suggestions in the comments, and perhaps come up with your own.

Caption: among my suggested ‘fossils we want to find’ are protobats (like the hypothetical examples shown at left, from Graham (2002)) and a good skeleton of the giant hominid Gigantopithecus blacki. This ilustration of a lower jaw is from Simons & Ettel's (1970) magazine article. Images: Graham (2002), Simons & Ettel (1970).
A habitually bipedal, large, non-human hominid. Whatever you think of all those stories, anecdotes and sightings about bigfoot, yeti, almas, orang-pendek, yowie and so on and on, the fact remains that the discovery of a large, bipedal non-human hominid – whether it be a pongine, hominine, or member of another hominid lineage – would be a huge deal. It would not just be one of the most newsworthy creatures to ever be discovered; it would also have enormous ramifications for our understanding of hominid evolution and potentially the human condition itself.

Caption: are crypto-hominids a cultural phenomenon more than a zoological one? I’ve argued for both possibilities at different times. Whatever… for the purposes of the article you’re reading now, I hope we can agree that the discovery of such an animal would be high on any hypothetical ‘wants’ list. Image: Darren Naish.
It would also – if relating to North America or northern Eurasia in particular – very likely have a significant impact on economy, land management and land use in those regions… or, you’d hope it would, anyway (who knows, given the current state of environmental protection in the USA). The hypothetical discovery of such an animal would also be regarded by many as one of the biggest ‘wins’ ever scored against ‘establishment science’, and thus could well be a bad thing (viz, “if scientists were wrong about this, what else could they be wrong about?”). And I’ll stop there before we dive into a rabbit-hole of conspiracy theories and coverups.
A big, flightless passerine. The majority of living bird species – over 60% of them – are passerines, or perching birds. This is the great group that includes crows, thrushes, warblers, finches, sparrows and so many others. For all their success, wide distribution and diversity, passerines are generally quite samey. There are no big, long-legged wading passerines, or heavy-bodied diving passerines or flightless running passerines, for example. Why this is so remains mysterious: passerines didn’t take to those niches because… well, they just didn’t. Does this mean that they couldn’t? As usual, we can come up with a few reasons as to why they were ‘constrained’ in evolutionary potential, but any one of those reasons could be overturned by some evolutionary deviant that refuses to pay attention to the rules.

Caption: passerine birds are diverse, to a degree… here’s just a sample of their diversity. This is part of a giant montage that’s being built for my in-prep textbook The Vertebrate Fossil Record. Image: Darren Naish.
And thus I submit that a particularly large, wholly flightless, cursorial passerine should make itself known to the world. It should be a record-holder as goes size, but not necessarily be that much bigger than the largest known passerines (like lyrebirds and ravens): I’m talking about a bird that weighs 3-5 kg and is thus similar in size to a large chicken. It should be a big, long-legged rail-babbler, quail-thrush or similar, and hence be a denizen of Wallacea or nearby.

Caption: Eupetes, the Malaysian rail-babbler. A hypothetical big, flightless passerine should be a close relative of this bird. Image: Francesco Verronesi, CC BY-SA 2.0 (original here).
A few recently extinct, island-dwelling passerines were flightless, so we do know that passerines have the evolutionary potential to follow this pathway. Such species (a bunting and a few New Zealand wrens… and possibly a few others) were all small (less than 40 g).
A western Asian giant salamander. Giant salamanders (cryptobranchids) are restricted today to eastern Asia (where Andrias occurs) and North America (where Cryptobranchus occurs). Hunting, human disturbance, habitat loss and deterioration, climate change and other issues are putting them into perilous decline, right at the same time as we’re discovering that some of them are species complexes. They were more widespread in the past than they are today, since fossils show that Andrias salamanders were widespread across Europe and Asia between about 28 and 2 million years ago.

Caption: an Asian giant salamander (Andrias) photographed in captivity. Record-holding specimens of Andrias can be 1.8 m long and exceed 60 kg, and some extinct species reached even larger sizes. Image: Markus Bühler.
While there are very good reasons for the decline and extinction of the animals in the areas concerned, some of the regions where they formerly occurred still have what look like suitable habitat today and are sparsely populated by people. Furthermore, extinct giant salamanders weren’t all denizens of fast-flowing, highly oxygenated streams like those inhabited by the modern populations. Some inhabited ponds and lakes. Ergo: I would really, really like there to be a west Asian cryptobranchid that comes from a habitat considered weird for the other living members of the group. And it doesn’t have to be a giant of 2 metres or more. A hellbender-sized species of 70 cm or so will do fine thank you very much.

Caption: some extinct cryptobranchids - this is Zdeněk Burian’s reconstruction of Andrias scheuchzeri - inhabited European ponds and lakes. I’ve previously criticised this image for showing the animal as terrestrial. Since then, the proposal has been made that some extinct cryptobranchids (albeit not A. scheuchzeri) were significantly more terrestrial than living species. Image: (c) Zdeněk Burian.
Again, this is an area of special interest to cryptozoologists, since there have been occasional suggestions that stories, engravings and such from western Asia might reflect folk knowledge of unusually big salamanders in the region. In reality, the images and stories concerned are super-ambiguous and more likely refer to otters and god knows what else.
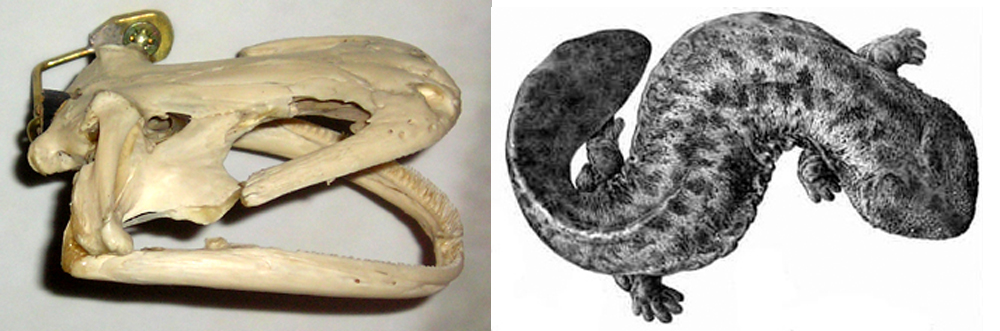
Caption: at left, an Andrias skull. Image: Darren Naish. At right: Japanese giant salamander (A. japonicus) illustration by Y. de Hoev from 1887. Image: Y. de Hoev, public domain (original here).
A living albanerpetontid. Everyone knows that there are three main groups of living amphibian: caecilians, salamanders and anurans (frogs and toads). But until (geologically) recently, there was a fourth group: the albanerpetontids, sometimes termed albies by those who work on them. Albanerpetontids were geographically widespread, their range including Eurasia, northern Africa and North America, and they were geologically long-lived too. The oldest are from the Middle Jurassic while the youngest are… well, we’ve known of Miocene fossil albanerpetontids for decades, have known of Pliocene specimens since 2005 (Venczel & Gardner 2005), and now know that at least one species persisted into the Pleistocene (Villa et al. 2018). The fact that their fossil record has been creeping towards the Recent means that the possibility of fossil and even extant Holocene specimens being discovered isn’t ridiculous, especially given the small size of these animals and hence tiny size of their bones.
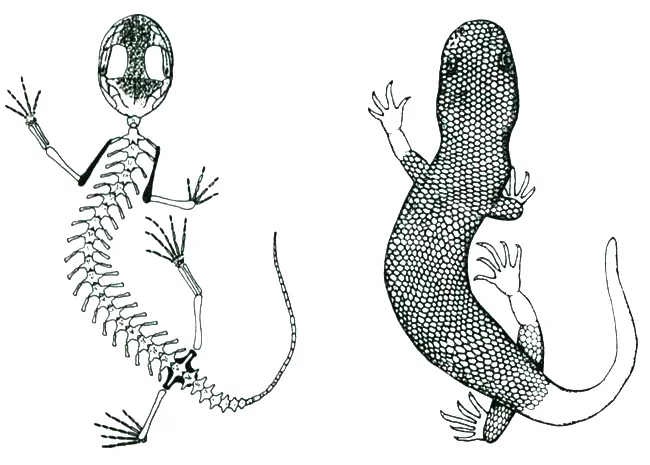
Caption: new salamander species are occasionally discovered in Europe and Asia even now. It would be amazing if an animal suspected to be a ‘new salamander’ one day turned out to be a living albanerpetontid. These reconstructions were published by McGowan & Evans (1995). They might have erred in implying that the scales would be externally visible as shown here; more likely is that they were concealed by epidermis, as in other scaly fossil amphibians. Image: McGowan & Evans (1995).
To be frank, a live albanerpetontid wouldn’t be a particularly spectacular animal: it would be a tiny, slim, salamander-like amphibian less than 10 cm long, and it wouldn’t be much fun to watch since it would spend most of its time hiding and burrowing in leaf litter. But among herp-nerds it would be a huge deal. Live albanerpetontids were scaly-skinned (though the scales were not necessarily visible externally), with eyelids, and with adaptations in the snout, skull-roof, neck and body shape linked to head-first burrowing (McGowan & Evans 1995).

Caption: an artistic reconstruction of a live albanerpetontid… produced for my in-prep The Vertebrate Fossil Record. Image: Darren Naish.
A Eurasian palaeognath. Palaeognaths are the big, flightless ratites (ostriches, emus and so on), the superficially gamebird-like, flight-capable tinamous, and their extinct relatives. A huge amount has been written about the evolutionary history and biogeography of these birds, since their distribution is curious and has resulted in all kinds of different models about how they might have spread around the world. I’ve written about this issue at length on previous occasion (the articles concerned being famous for generating the longest-ever comment threads in the history of TetZoo… though all of this is mostly wasted now, what with SciAm’s paywalling of the site, sigh). Living palaeognaths are absent from Eurasia, despite the former present in the region of ancient, flight-capable Paleogene taxa, extinct ostriches and others.

Caption: as this map shows, modern palaeognaths occurred everywhere until recently (except Antarctica) with the exception of northern North America and the cooler parts of Eurasia. Extinctions across Eurasia, Madagascar and New Zealand of course saw the disappearance of various members of the group. Image: Darren Naish.
The fact that Paleogene Europe was home to many bird groups that no longer occur there but are now denizens of tropical regions elsewhere leads me to hope for a living palaeognath – a tinamou- or bustard-sized species – that descends directly from archaic Paleogene taxa and now lives in the Asian tropics. It should be a cryptic generalist with barred plumage and a mid-length bill and a reduced flight ability.
A gigantic, predatory, limbed amphisbaenian. Regular readers of TetZoo might know that I really like amphisbaenians: the mostly limbless, bullet-headed ‘worm lizards’ of the American tropics, Africa, and parts of southern Europe and western Asia. Amphisbaenian evolutionary history and biogeography has become increasingly complex in recent years as we’ve learnt a bunch of new stuff about their fossil history, genetics and anatomy. Among the weirdest of amphisbaenians are the ajolotes (or bipedids), the only extant group to possess limbs. These limbs are not small stumps or flaps (as they are in some other near-limbless, serpentine squamates) but well-developed, clawed forelimbs. According to some phylogenetic models, ajolotes are not the sister-group to limbless amphisbaenians but deeply nested within the limbless clade (Conrad 2008, Videl et al. 2008), in which case their limbedness – if you will – perhaps evolved from limbless ancestors. Add to this the fact that some amphisbaenians are robust-jawed, short-faced predators of vertebrates that ambush prey from beneath the surface and bite chunks from the bodies of surface-dwelling mammals and reptiles.
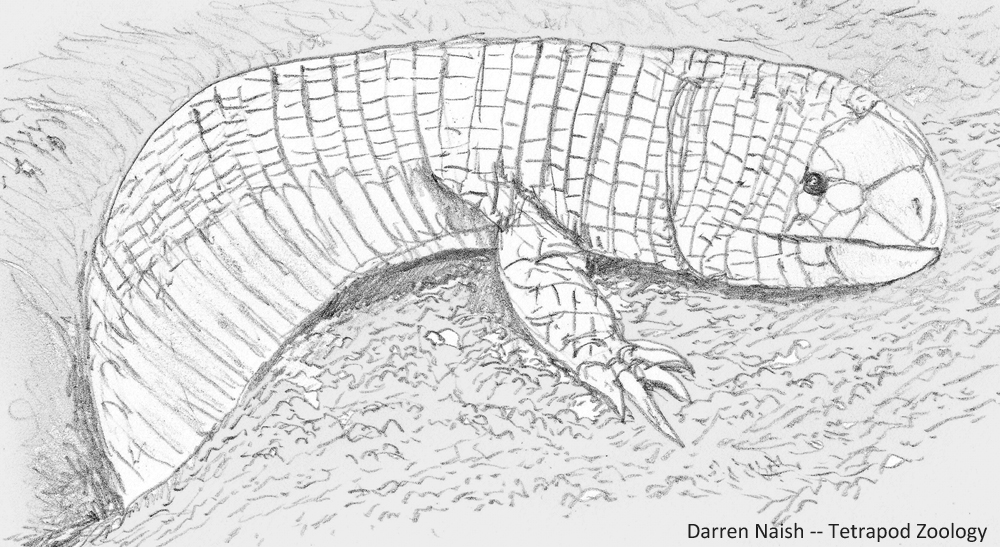
Caption: Bipes, an ajolote of Mexico (they might occur in parts of the USA as well). Three extant species are recognised. Image: Darren Naish.
So then… where oh where are the giant, limbed, robust-skulled, vertebrate-eating amphisbaenians? By ‘giant’, I am not talking about a graboid-sized monster of several metres (though that would be nice), but a more reasonable animal of a mere 1.5 metres or so. Easily the stuff of nightmares. They could inhabit warm regions of any continent.

Caption: Carl Gans’s illustration of a burrowing ajolote, showing how the large, well-clawed forelimbs function in propulsion. This is clearly a Five-toed worm lizard Bipes biporus; the other extant species have four and three digits, respectively. Image: Gans (1974).
Is there any reason to think that gigantic, predatory, limbed amphisbaenians might actually exist and await discovery? Nope. But I wish it were so. Regular readers might recognise that such creatures are denizens of the alternative-timeline Earth of the Squamozoic, but I’m sure that that’s coincidental.
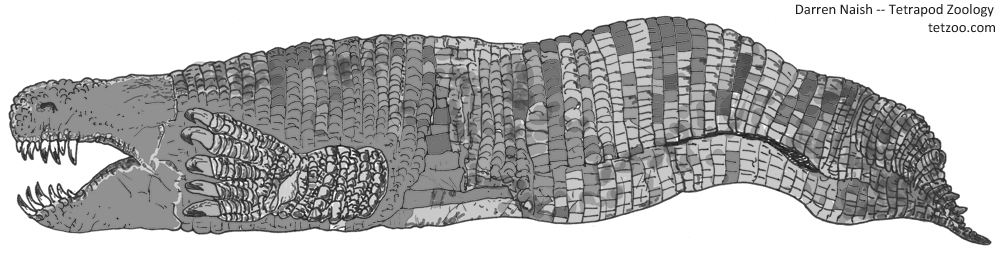
Caption: what would a gigantic, predatory, limbed amphisbaenian look like? Like this, of course. Image: Darren Naish.
An African or west Eurasian, long-beaked river dolphin. On several occasions within the history of odontocete cetaceans (‘toothed whales’), lineages have moved into brackish and estuarine environments, and eventually made the transition to committed freshwater life. There are the Asian Platanista species, the recently extinct Lipotes of China, and the tropical American Inia species. Once united within Platanistoidea and thought to be close kin, we know today that these animals represent at least three separate transitions to the freshwater environment (the term Platanistoidea is now restricted to the Platanista lineage alone). In addition, members of other groups – I’m thinking of the delphinid Orcaella – occur in rivers within parts of their range. There’s also a fossil beaked whale that might be indicative of freshwater specialisation in yet another odontocete group (Mead 1975).

Caption: river dolphins are pretty special looking. This is a Ganges river dolphin (Platanista gangetica). Image: Zahangir Alom / Marine Mammal Commission / National Oceanic and Atmospheric Administration, public domain (original here).
In view of all this, why aren’t there river-dwelling dolphins in Africa, Europe or western Asia? Again, the answer seems to be… there just aren’t. A few fossil taxa suggest that such animals might have evolved if things had gone another way (there are fossil platanistoids from the Caucasus, for example). But I humbly submit that the great river systems of tropical Africa, the Tigris-Euphrates system of western Asia and the Danube, Po, Ebro, Dniester and others of southern Europe would be much improved if only we knew of their endemic riverine dolphins. I’m talking about a true riverine specialist, convergent with Inia and Platanista, with a long beak, spike-like teeth, reduced eyesight, the works. And if you want to play fast and loose with antiquarian literature and anecdote, there are references in the literature to ‘river dolphins’ in the Nile and there are even one or two eyewitness accounts from central Europe that describe long-beaked ‘dolphins’ seen in rivers and lakes.

Caption: if there are extant west Eurasian or African river dolphins, they should look like this. This is a hypothetical species, modelled on the American Inia and Asian Platanista. Image: Darren Naish.
An endoparasitic tetrapod. Tetrapods have become parasites on several occasions. Vampire bats are parasites of birds and mammals, and it’s even been argued that some blood- and milk-eating human populations can be considered parasites of the mammals they rely on (though the mammals concerned are domesticated, so it’s complicated). Elsewhere among vertebrates, everyone knows about the parasitic catfishes that invade the gills of other actinopterygian fishes and even the urethras of mammals; less familiar is the fact that other actinopterygians can, on rare occasion, become trapped inside the bodies of other vertebrates and then make a successful living. Yes, you read that right. I have in mind the case where two Snubnosed or Pugnose eels Simenchelys parasitica were discovered living inside the heart of a mako shark (Caira et al. 1997; see also Eagderi et al. 2016). This eel is not – despite its name* – ordinarily an internal parasite: this was a case of facultative endoparasitism!

Caption: at left, a snubnosed eel found living inside the heart of a shark. Eels are not tetrapods, it’s true. But here’s evidence that aquatic vertebrates can become endoparasites. Image: Caira et al. (1997). At right: an aquatic typhlonectid caecilian. Surely it’s only a matter of time before we discover an endoparasitic one of those as well. Image: Neil Phillips.
There are all kinds of reasons why a tetrapod couldn’t become an endoparasite, respiration being high on the list. A hypothetical endoparasitic tetrapod would have to be small, with remarkable tolerance of unusual chemical and thermal conditions, with low oxygen requirements, and most likely with the ability to respire cutaneously or via gills. In other words, it should be the world’s weirdest caecilian. As if caecilians aren’t weird enough, I’d love there to be small, endoparasitic caecilians. Given that some caecilians are already aquatic gill-breathers that will consume the tissues of fish (exhibit A: the sequence from River Monsters where Jeremy Wade discovers swarming typhlonectid caecilians in the carcass of a large fish), I predict these animals to be aquatic, South American species that parasitise actinopterygians and aquatic mammals, like Inia the river dolphin.
* Snubnosed eels were given the name ‘parasitica’ because they opportunistically latch on to the bodies of larger fish and eat away at the flesh. They were not thought to ever be proper internal parasites prior to 1992.
And that’s where I’ll stop for now. I actually came up with a list containing numerous additional ‘wish list’ animals but time is against me. Maybe I’ll cover them in another article. Whatever, this was all a bit of fun and I hope you enjoyed it.
For TetZoo articles relevant to the issues covered here, see…
Surreal caecilians part I: tentacles and protrusible eyes, January 2008
Surreal caecilians part II: pass mum’s skin, hold the mayo, January 2008
Close up to Andrias, despite the smell and the teeth, December 2010
Welcome to the Squamozoic!, April 2013
If Bigfoot Were Real, June 2016
PS I’m going to stop linking to the SciAm run of TetZoo articles soon, because I cannot access them at all and they’re now all but useless. They all need to be relocated to an open-access site.
Refs - -
Caira, J. N., Benz, G. W., Borucinska, J. & Kohler, N. E. 1997. Pugnose eels, Simenchelys parasiticus (Synaphobranchidae) from the heart of a shortfin mako, Isurus oxyrinchus (Lamnidae). Environmental Biology of Fishes 49, 139-144.
Conrad, J. 2008. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bulletin of the American Museum of Natural History 310, 1-182.
Eagderi, S., Christiaens, J., Boone, M., Jacobs, P. & Adriaens, D. 2016 Functional morphology of the feeding apparatus in Simenchelys parasitica (Simenchelyinae: Synaphobranchidae), an alleged parasitic eel. Copeia 104, 421-439.
Graham, G. L. 2002. Bats of the World. St. Martin’s Press, New York.
Mackal, R. P. 1987. A Living Dinosaur? In Search of Mokele-Mbembe. E. J. Brill, Leiden.
McGowan, G. J. & Evans, S. E. 1995. Albanerpetontid amphibians from the Cretaceous of Spain. Nature 373, 143-145.
Mead, J. G. 1975. A fossil beaked whale (Cetacea: Ziphiidae) from the Miocene of Kenya. Journal of Paleontology 49, 745-751.
Naish, D. 2017. Hunting Monsters: Cryptozoology and the Reality Behind the Myths. Arcturus, London.
Simons, E. L. & Ettel, P. C. 1970. Gigantopithecus. Scientific American 222 (1), 77-84.
Venczel, M. & Gardner, J. D. 2005. The geologically youngest albanerpetontid amphibian, from the Lower Pliocene of Hungary. Palaeontology 48, 1273-1300.
Vidal, N., Azvolinsky, A., Cruaud, C. & Hedges, S. B. 2008. Origin of tropical American burrowing reptiles by transatlantic rafting. Biology Letters 4, 115-118.
Villa, A., Blain, H.-A. & Delfino, M. 2018. The Early Pleistocene herpetofauna of Rivoli Veronese (Northern Italy) as evidence for humid and forested glacial phases in the Gelasian of Southern Alps. Palaeogeography, Palaeoclimatology, Palaeoecology 490, 393-403.



