Back in July 2019, myself and a bunch of friends stepped aboard the Pont-Aven for several days of sea-watching in the Bay of Biscay. We were to travel from Plymouth (UK) to Santander (Spain), the event being organised by ORCA, a charity that monitors whales and uses the data for conservation purposes (they’re here on Twitter). ORCA uses cruise liners, ferries and other vehicles as whale-watching platforms. Nigel Marven was a special guest on our trip and it was great to catch up with him.

Caption: our vessel of choice - the Pont-Aven - at port in Santander, Spain. I cannot tell you how much trouble I went to to get to this ship before departure time. I very nearly didn’t make it. Image: Darren Naish.

Caption: the man, the legend; Nigel Marven.
The purpose, of course, was to see whales. The weather was outstandingly good (meaning that I got burnt), but so was the whale watching: I’m pleased to say that we saw literally hundreds of animals of seven or eight species, as you can see from the photos below. My own photos are not great since my camera isn’t exactly the best for fast-moving, far-away animals like whales, so those you see here were mostly taken by my trusty pal Alex Srdic (who’s here on Instagram and here on Twitter). Thanks, Alex.

Caption: several cetaceans have extremely complex markings allowing them to be identified to species and even population. Individuals can be recognised on the basis of their markings too. Image: Alex Srdic.
The Bay of Biscay is a world-famous whale-watching hotspot, famous in particular for Cuvier’s beaked whales Ziphius cavirostris and Sperm whale Physeter macrocephalus. Dolphins of several species are a frequent sight too, as are rorquals of a few species, Harbour porpoise Phocoena phocoena and pilot whales. A very lucky whale-watcher might get to see Blue whale Balaenoptera musculus, Killer whale Orcinus orca or True’s beaked whale Mesoplodon mirus. In fact, something like 30 species have been recorded in the region. This is phenomenal and mean that it’s theoretically possible for several species of some of the most elusive whale groups – like beaked whales and globicephaline dolphins – to be seen within days or weeks of each other.

Caption: in good weather, the blow of a big whale (like a Fin whale - as here - or a Sperm whale) is visible from great distance, and in the case of these two species can be diagnostic. Image: Alex Srdic.

Caption: a dynamic leap by a Striped dolphin. Dolphins of some species appear to be attracted to ships and even to deliberately show off when they get close to them. Image: Alex Srdic.
Why is the Bay of Biscay so good for whales? It’s mostly because the topography is complex, combining large, shallow shelf regions, steep sections of shelf edge – sometimes with impressive slopes and deep, enormous rocky canyons twice as big as the Grand Canyon – and a deep abyssal plain section (Carwardine 2016). Depth varies from 1.7 to over 4.7 km. This variation – combined with the overall productivity of the region and its position relative to the Atlantic and English Channel – means that there’s the chance to see continental shelf species (like porpoises), those that use deep canyons and other shelf-edge habitats (like beaked whales) and true oceanic deep-divers that forage in the deepest waters (like sperm whales).

Caption: back and dorsal fin of a Fin whale, remnants of the blow still hanging in the air. Image: Alex Srdic.
As it happens, we were extraordinarily lucky. Fin whales B. physalus are regular animals of the area, and we had amazing, relatively close views of them (by ‘close’, I mean perhaps 30 m from the ship, not alongside the vessel). Fin whales – the second largest extant animal after the Blue – have a blow that’s visible on the horizon and is about 8 m tall. The blow hangs in the air for a surprising time. One of the most remarkable things about the Fin whale is its asymmetrical pigmentation: the right side of the face is marked with a large pale area, as is the right side’s baleen. There are some old TetZoo articles on what this might mean and how it might function – see the links below.

Caption: excellent view of the splashguard - the conical structure surrounding and ahead of the blowholes - and paired blowholes of a surfacing Fin whale. Despite its name, the dorsal fin of the Fin whale is smaller and blunter than that of some other rorquals. Image: Alex Srdic.
Two coastal species were seen early on in our trip: Harbour porpoise and Common bottlenose dolphins Tursiops truncatus, though I don’t have good photos of either. The majority of dolphins seen on our trip (as is typical for Biscay whale watching) were Short-beaked common dolphin Delphinus delphis, which were sometimes seen in groups of more than ten. Their distinctive hourglass markings are always visible when they leap – which they often do, sometimes while immediately adjacent to a ship – and we also got to see calves on one or two occasions.

Caption: here’s the whole-body view of the common dolphin shown in detail above. This individual only has one stripe extending from the beak to the flipper, with a large pale area separating the eye and flipper. Different configurations are present in different populations. Image: Alex Srdic.

Caption: as the light begins to fade during the later part of the day, a group of Short-beaked common dolphin carve through a surging wave. Note the calf close to the adult at upper right. Image: Alex Srdic.
We also had excellent views of Striped dolphin Stenella coeruleoalba. They behaved in characteristic acrobatic fashion, leaping high out of the water, making impressive splashes and jumping in the ship’s wake. They typically make a lot more disturbance at the water’s surface than do common dolphins, creating great bursts of spray and rooster-tail patterns when they leap and surge. Striped dolphins are near-globally distributed. They’ve been the source of discussion lately since it’s recently been shown that the Clymene dolphin S. clymene is a naturally occurring hybrid between this species and the Spinner S. longirostris (Amaral et al. 2014).

Caption: we had many excellent views of high-leaping Striped dolphin. Note how much spray and splashing is associated with the leaping of this species. Image: Alex Srdic.
Finally as goes dolphins, we also saw pilot whales, identified on the basis of their black colouration and strongly backswept dorsal fins. These were most likely Long-finned pilots Globicephala melas (it’s more typical of temperate and cold waters than the Short-finned G. macrorhynchus) but we didn’t see any of the key features that allow the two species to be distinguished, and none of our photos are good enough to warrant sharing. A mysterious whale was seen among the pilot whales. It seemed to be very dark and with a short, blunt-tipped, parallel-sided but only weakly curved dorsal fin; I don’t think that its head was seen but I had the impression that it was a shallower-bodied animal than the pilot whales. Several different views were offered on its identity with the most likely (on the basis of dorsal fin shape) being that it was perhaps a False killer whale Pseudorca crassidens. That’s not tremendously likely but not impossible.
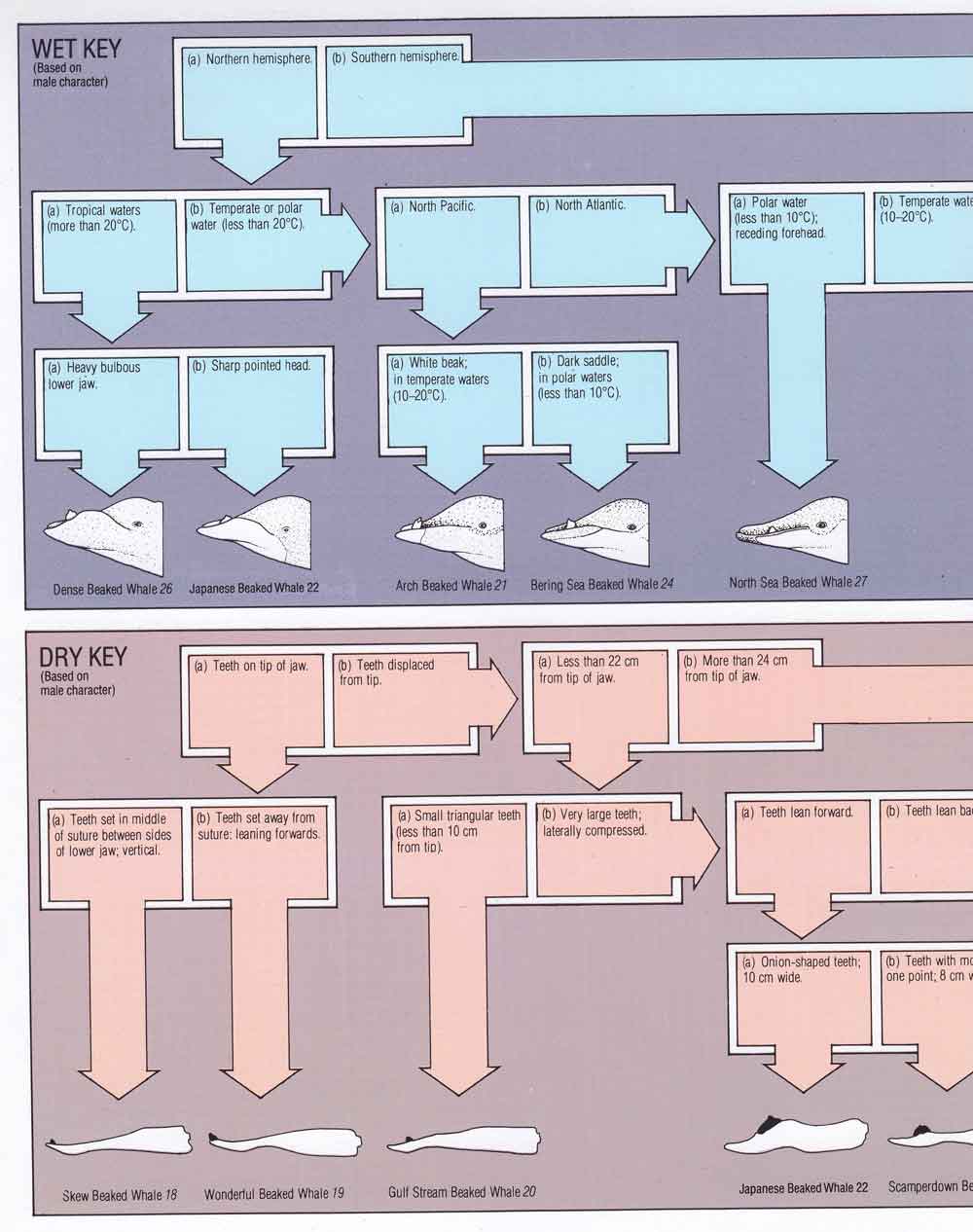
The whale most famously associated with the Bay of Biscay is Cuvier’s beaked whale, seen so frequently in the area that it’s regarded as the premier location for sightings of this species, worldwide. I don’t know if you’re guaranteed a sighting of a Cuvier’s while there, but – whatever – we were lucky, since we saw nearly 20 of them, ranging from smooth, clean-bodied youngsters to heavily scarred males.

Caption: Cuvier’s beaked whale, seen relatively close to the ship. Image: Alex Srdic.

Caption: heavily scarred Cuvier’s beaked whale, seen at distance and only briefly. We didn’t see any other individuals with scarring as impressive as this. Image: Alex Srdic.
Some individuals have markedly pale heads sharply demarcated from the rest of the body, others do not. On occasion, one or two individuals were close enough to the ship that I was able to get a half-decent shot with my mobile phone. Each sighting was a huge thrill. While we were oh so lucky as goes Cuvier’s, we didn’t see sperm whale, alas. We also saw Northern minke B. acutorostrata on perhaps two occasions, though again I don’t have any good photos.

Caption: another plus… amazing sunsets, and sunrises too. Image: Darren Naish.
Finally, we didn’t just see whales. The same route is also great for seabirds, and we also saw such fishes as tunas and sunfishes. As much as I’d like to start talking about the birds, I’m out of time. Anyway – the trip was excellent: rewarding, fun, and educational. I’ll definitely be doing it again. You should consider supporting ORCA and their work as well.
Cetaceans have been covered at length on TetZoo before - mostly at ver 2 and ver 3 - but these articles are now all but useless since all of their images have been removed (and/or they’re paywalled, thanks SciAm). Over time, I aim to build up a large number of cetacean-themed articles here at ver 4.
A 6 ton model, and a baby that puts on 90 kg a day: rorquals part I, October 2006
From cigar to elongated, bloated tadpole: rorquals part II, October 2006
Lunging is expensive, jaws can be noisy, and what’s with the asymmetry? Rorquals part III, October 2006
On identifying a dolphin skull, July 2008 (all images now missing)
Seriously frickin' weird cetacean skulls: Kogia, shark-mouthed horror, July 2008 (all images now missing)
Scaphokogia!, July 2008 (all images now missing)
Cetacean Heresies: How the Chromatic Truthometer Busts the Monochromatic Paradigm, April 2015 (but now lacking all images)
Refs - -