Let’s experiment with camera trapping!
Tet Zoo Reviews Zoos: Tokyo’s Ueno Zoo
New Species Round-up for 2023, Part 2
In the previous article we looked briefly at those new amphibian and mammal species named during 2023. This time we skip ahead to reptiles… including birds because – yes – birds are reptiles in the phylogenetic sense)….
Birdwatching in Tajikistan, Part 2
Birdwatching in Tajikistan, Part 1
Alan Feduccia’s Romancing the Birds and Dinosaurs: Forays in Postmodern Paleontology
Laysan Ducks: Not as Degenerate as People Used to Think
The Great Florida Cassowary Relocation Event of 2023
Controversies in Ratite and Tinamou Evolution (Part I)
Surprising Diet of the House Sparrow
Otidiphaps the Pheasant-Pigeons
Inside the Cassowary's Casque, Updated for 2022
Chiffchaffs and What Are 'Old World Warblers' Anyway?
Pete Dunne and Kevin T. Karlson’s Gulls Simplified: A Comparative Approach to Identification
Painted-Snipes
Kirk W. Johnson’s 2018 The Feather Thief, a Review
Why the World Has to Ignore David Peters and ReptileEvolution.com
The Dodo in Life
You’re already highly familiar with the Dodo Raphus cucullatus, and images of what this animal looked like in life are present in a vast number of books, popular sources and museum displays. What might not be so familiar to you is that the Dodo’s life appearance has been the subject of long-standing debate, that many familiar ideas about its appearance are very likely wrong, and that – even today – we’re uncertain about several details.
Suburban Birdwatching in Queensland, Australia
I recently spent time in Australia, and specifically in Brisbane, Queensland (this was for the 79th Society of Vertebrate Paleontology annual meeting), and, while there, I got to see a pretty good selection of birds. I photographed as many as I could, and in this article I’m going to talk about them. All the birds I’m going to discuss here were encountered in urban and suburban settings in close proximity to people, and none are especially exotic or obscure. But they were entirely novel to me, and I was hugely excited to encounter them in the wild. I hope you’ll enjoy reading about them.

Caption: Dusky moorhen, Australian white ibis and Little black cormorant captured in the same one shot (at left). At right: a wonderful Torresian crow. Images: Darren Naish.
One more bit of preamble: if I had the time and foresight, I could have organised a special birdwatching tour, in which case I could have been taken to various specific locations at which I might have seen a rather more impressive list of Australian birds not ordinarily encountered by mere chance. But… I didn’t. I wish I had.

Caption: if you’re truly interested in animals, you’re interested in invasives, aliens and introductions, and there’s no shame in paying attention to them or studying them. So go ahead and photograph that feral pigeon. Image: Darren Naish.
My very first Australian bird sighting was of Welcome swallows Hirundo neoxena, seen flying around the outside windows of Perth Airport. I was to see this species on several subsequent occasions (it occurs across virtually the whole of Australia except for some north-central sections), but no photos, sorry. My second species: the feral Rock dove or Rock pigeon Columba livia! I don’t care what anyone says: I always take time to look at feral pigeons, and one thing you notice is that populations differ from place to place, mostly because they descend from different founding populations of escapees or released birds. Brisbane pigeons were especially dark relative to the majority of familiar urban pigeons in Europe, and on the large size too. Brisbane: make of that what you will.

Caption: my first bin chicken. This individual spent time probing into the gap between the pavement and a retaining wall, searching for arthropods. Image: Darren Naish.
My second Aussie bird was something far more exotic (to me): an ibis, and specifically an Australian white ibis Threskiornis molucca. This species was included within T. aethiopica – the species currently referred to as the African sacred ibis – until the mid-2000s, and some authors still regard the two as conspecific. The Black-headed ibis, Black-necked ibis, Oriental white ibis or Indian white ibis T. melanocephalus is part of this complex too (all three are regarded as part of the same superspecies). Apparently, there are Australian people who still refer to T. molucca as the Sacred ibis but --- as any Australian will tell you within 0.5 seconds of you expressing interest in this bird, it’s not ‘the Australian white ibis’ to the vast majority of urban Australians but ‘the bin chicken’: an animal that people associate with rubbish, waste food and urban filth in general. I get the impression that Aussies love to hate the ibis the same way people also dislike urban gulls and pigeons. Hey idiots... the shitty trash the bird is eating is, like, YOUR trash and YOU put it there.

Caption: Australian white ibis out and about in town. Image: Darren Naish.
Australian white ibises seem to be both abundant in Brisbane and extremely bold. Large groups are present in the parks throughout the city, groups can be seen nesting at the tops of palms and other trees in the parks, and individual birds can be seen walking around in crowded pedestrianised areas and even right into cafes and restaurants… though they aren’t exactly welcome in such places.

Caption: large numbers of ibises are readily visible in parks and other green places in Brisbane. The birds stand or sit on the grass and rest in trees. The first individual I saw (the one probing in the paved area, shown above) was initially sat on the grass, chest against the ground. Image: Darren Naish.

Caption: aaaand… even more ibises. Image: Darren Naish.
Go near the water most places in the world, and you’ll very likely see members of the duck, goose and swan family Anatidae, and such it is in Australia. I saw three species, all new to me, all in close proximity to the ponds in Brisbane’s Botanical Gardens. We’ll start with the Australian wood duck, Maned duck or Maned goose Chenonetta jubata, a heavily terrestrial, pan-Australian, cavity-nesting duck that might be a member of the shelduck clade Tadorninae. The birds I saw were asleep (or pretending to be asleep) and thus standing and sitting still with their eyes closed.

Caption: at left, two Australian wood ducks. resting or sleeping. At right: a Hardhead heads across the water towards me. Presumably, it (and its partner) was used to being fed by people. Image: Darren Naish.
Pacific black ducks Anas superciliosa – PBDs in birding vernacular – were also seen; this is an especially handsomely marked member of the mallard complex, the different forms of which are quite variable across its southwest Pacific/Australasia range. Another first for me was Hardhead or White-eyed duck Aythya australis, a Pacific/Australasian member of the scaup and pochard lineage (Aythyini).

Caption: Pacific black duck. Of the three recognised subspecies, the Australian one is A. superciliosa rogersi. This subspecies also occurs in Indonesis and New Guinea. Image: Darren Naish.
My biggest surprise at the ponds, however, was an Australian darter or anhinga Anhinga novaehollandiae (here I’m following recent studies in recognising this as a distinct species from African and Asian anhingas), which I watched for some time as it foraged and hunted close to the water’s edge. On occasion, it would disappear entirely from view before stretching its long, slender neck out of water while swallowing a fish. And on other occasions, its long tail could be seen part emerging above the surface while the bird was part swimming, part floating, just beneath the surface. I’ve never seen a live anhinga before. This one was smaller than I expected.

Caption: an anhinga surfaces and swallows what I presume is a fish. Image: Darren Naish.

Caption: more swimming anhinga action. Image: Darren Naish.

Caption: Little pied cormorant, foraging in a shallow pool with abundant algae and other aquatic plants. Its appearances at the surface were really brief compared to the other cormorants I’ve watched, and this is the only photo that clearly shows its head and bill. Image: Darren Naish.
Two cormorant species were also present at the ponds: Little black cormorant Phalacrocorax sulcirostris (see photo at the top of the article) and Little pied cormorant P. melanoleucos (or Microcarbo melanoleucos, if you follow Siegel-Causey’s (1988) taxonomy). The one individual of the latter I saw was diving for food in a heavily vegetated pool. A group of three Dusky moorhens Gallinula tenebrosa were paying close attention to its activities, and would circle and peck at the surface every time the cormorant would dive. I assume that they were interested in bits of vegetation and small animals brought to the surface by the cormorant’s activities. Indeed, numerous small fish and tadpoles were present in the pools. The tadpoles looked like those of a toad and I presume they were Cane toad Rhinella marina larvae, indeed I know that Cane toads were present since I found two dead ones in the park grounds.

Caption: Dusky moorhens search for food in an area where the water is being disturbed by a swimming Little pied cormorant. Image: Darren Naish.
Dusky moorhens were frequently encountered and easily approached. As suggested by the name, they looked darker than the Eurasian moorhens G. chloropus I know well, and they appeared larger and chunkier too. I watched them foraging on lawns, and a bit of squabbling, chasing, fighting and mating was seen as well.

Caption: Silver gulls look similar to Northern Hemisphere species like the Black-headed gull Chroicocephalus ridibundus, and there’s a reason for that. Image: Darren Naish.
Finally on birds associated with watery places, I also visited an artificial beach close to the Brisbane River where there were numerous Silver gulls Chroicocephalus novaehollandiae. This is Australia’s most widespread gull and occurs around the country’s entire coast as well as in many inland locations. The Silver gull is surrounded in phylogeny by species that have grey, black or brown heads (Chu 1998) and is thus generally agreed to be part of the Chroicocephalus group (and is thus not part of Larus in the new, restrictive sense). At least a few other Chroicocephalus species are like the Silver gull in being white-headed too, so it might be that a transition from a dark head back to a white one happened a few times in this group. The Silver gull is a mid-sized, slender-billed gull.

Caption: Masked lapwing, or Black-shouldered lapwing if you’re so inclined. Note that the carpometacarpal spurs aren’t readily visible here. Image: Darren Naish.
I also saw a single plover, specifically a Masked lapwing Vanellus miles… and, more specifically, the black-shoulder form V. m. novaehollandiae (jeez, colonial Europeans: couldn’t you be a bit more creative with the scientific names?) sometimes recognised as the distinct species V. novaehollandiae, the Black-shouldered lapwing. This bird was wandering around on a lawn close to a restaurant. Lapwings – technically called vanellines – are conventionally included within Charadriidae, the plover family. Lapwings are interesting in that they possess large spurs on the carpometacarpus, which are typically (but not always) visible when the wings are closed. These are used in defence against predators (like cats, dogs and corvids) and also in intraspecific fights. There’s an apparent common folk belief that the spur is venomous! Here’s your regular reminder that TetZoo ver 3 includes two fairly comprehensive articles on the spurs, claws and clubs present on the wings of birds (part 1 here, part 2 here; I’ve linked to wayback machine versions as they include the illustrations).

Caption: a Laughing kookaburra in the wild, perched and vigilant. Image: Darren Naish.
I was surprised (hey, I’m from Europe) to see a Laughing kookaburra Dacelo novaeguineae in an urban park, sat on top of a streetlight and also in flight in the same park. I gather than kookaburras are pretty typical park and garden birds in parts of Australia, well known for their habit of grabbing food from tables and barbeques and such. Most of you will know that kookaburras are especially big kingfishers, closely related to Halcyon kingfishers and kin, and that the bulk of kingfisher diversity exists in the islands and coastal regions of Australasia. Kingfishers have conventionally all been included within the same one family (Alcedinidae), and this is still a popular view. However, the idea that lineages within the group are ‘distinct enough’ that several kingfisher families should be recognised is also popular, in which case kookaburras and their close kin belong within Halcyonidae (previously known, incorrectly, as Dacelonidae; Sibley & Ahlquist 1990). Miners (more on them in a minute) are not fans of kookaburras and a few individuals took time to mob the individual sat on top of the streetlight.

Caption: a Noisy miner (at left) arrives to harass a Laughing kookaburra. Image: Darren Naish.
I was also pleased to see Rainbow lorikeets Trichoglossus moluccanus at a few places, though none of my photos are at all good. Lorikeets mostly eat pollen and nectar and their specialised brush-like tongues explain their generic name – Trichoglossus means ‘hair tongue’. I actually saw parrots of a few other species as well, but they were right at the top of really tall trees and mostly seen in silhouette, so I never saw any of the detail that might allow them to be identified.

Caption: Rainbow lorikeet at the top of a palm tree. The green collar confirms the identification, since similar lorikeet species (like the Red-collared lorikeet T. rubritorquis) lack this feature. Image: Darren Naish.
As everyone who knows anything about birds knows, the majority of living bird species (over 60% of them) are passerines (or perching birds), and most of the Australian species I saw were members of this enormous group. I didn’t see any feral European starlings or sparrows, or indeed any introduced European species at all, and nor did I see any small endemic Australian endemics, like fairywrens, pardalotes or sunbirds.

Caption: a Noisy miner, eating parts of a Mexican burrito on a restaurant table. Image: Darren Naish.
We’ll start with Noisy miners Manorina melanocephala. These are mostly grey, thrush-sized passerines – happy to take to the ground as well as forage in trees and shrubs – that have a robust bill and naked yellow patch of skin behind the eye. They’re bold and look to have made the full transition to life in urban settings. I saw them jump around on tables and eat food from plates just moments after people had left, and I also saw them waiting for dropped food scraps right next to a person sat on the grass. In one case, a man eating his lunch while sat in the park was literally slapping the birds out of the way.

Caption: Common or India mynas are one of the most important alien birds in Australia (‘important’ in economic and ecological terms). Note that this individual is afflicted by a problem most commonly associated with urban pigeons: mangled toes and missing toe segments (the hallux claw is missing on the left foot). Image: Darren Naish.
The fact that these birds are called ‘miners’ is a bit confusing, since this makes it sound as if they’re ‘mynas’ (also spelt ‘mynahs’); that is, members of the starling and myna family Sturnidae. But they’re not, they’re honeyeaters (Meliphagidae). There are, incidentally, proper mynas in Australia: namely, the introduced Common or Indian myna Acridotheres tristis from tropical southern Asia. I saw some of these as well (I also saw them while on the journey home, in Singapore). Back to honeyeaters: Australia is the land of honeyeaters, the country being home to about half of the c 190 extant species. For all that I only saw one other meliphagid species: the Blue-faced honeyeater Entomyzon cyanotis.

Caption: a Blue-faced honeyeater, photographed at distance while it was on the top of a building. The area of blue facial skin is variable in these birds; this individual seemed to have only a small amount of blue. The skin is green in juveniles. Image: Darren Naish.
Moving on, Australia is also home to a really exciting assortment of sometimes large, omnivorous and predatory, superficially crow-like passerines, namely butcherbirds, currawongs and Australian magpies. I bet most people assume that these are corvids, but they aren’t: they’re conventionally allied within the family Cracticidae but a close relationship with woodswallows and kin means that the best course of action might be to include them within Artamidae, the woodswallow family. These birds are part of Corvoidea – the large passerine clade that includes shrikes, vireos, birds-of-paradise and corvids proper – but they’re some distance away from corvids, instead belonging to Malaconotoidea (also written Malaconotidea), a group that includes African bushshrikes and vangas (Cracraft et al. 2004, Cracraft 2014, Selvatti et al. 2015).

Caption: a Grey butcherbird (at left) waits for food in a park; to the right are a group of Noisy miners. Image: Darren Naish.
I saw a single Grey butcherbird Cracticus torquatus, hanging out in a park in close proximity to a group of miners. It was part of the group mentioned above, seen waiting near a person eating their lunch.

Caption: a (slightly blurry) Pied currawong caught in the middle of a run, just prior to takeoff. Image: Darren Naish.

Caption: Pied currawong from below. There are supposed to be three currawong species, but there are so many local variants, intermediate grades and island endemics that the story is quite difficult to resolve. Images: Darren Naish.
Pied currawong Strepera graculina were seen in trees, on tall buildings and structures attached to buildings, and on the ground. A pair were actually nesting on metal support structures just outside the conference venue but their nest was so high up and far from the ground that they were constantly at the edge of my camera’s range. Anyway, some of my photos aren’t terrible. And I did see Australian magpies on several occasions, but it was always while I was in cars, away from the city, and I never had the chance to get a photo.

Caption: Torresian crow, what an excellent bird. Image: Darren Naish.
Finally, Australia is also home to corvids proper. Australian crows are weird. To a European eye, they don’t look quite right – as if they might not be crows proper (which they are) – and yet they all look about enough alike that they could conceivably be close relatives (which they are: Jønsson et al. 2012). I saw members of two species, the first being a beautifully glossy, iridescent bird, about similar in size to typical Corvus crows like the Eurasian Carrion crow C. corone, and with a prominent pale iris. This was the Torresian crow C. orru, I assume named for the Torres Strait (the stretch of water between New Guinea and Australia). I saw this species both as a singleton and in a large group, often right in the middle of town.

Caption: more Torresian crows, both in a group, in a tree, and foraging on the ground in an urban area. Images: Darren Naish.

Caption: Torresian crows and flowers. I like the artiness of this photo. Image: Darren Naish.
The second crow species I saw was the larger, bulkier Australian raven C. coronoides. I saw a group of three of these, walking around together on a lawn. The large group of Torresian crows I saw appeared to be gathered together because they were unhappy about the proximity of these ravens, and they were peering in the ravens’ direction while noisily calling. I wanted to watch all of these corvids for longer, but my time was short and I had to run.

Caption: a pair of Australian ravens forage on a lawn, to the displeasure of nearby Torresian crows. Image: Darren Naish.
And that brings things to an end. There’s a vast amount of stuff that I never saw, much of it living in very close proximity to the part of Brisbane in which I was staying. But it wasn’t to be. One day I’ll visit Australia again, and I hope for better luck, more time, and more experience of the region’s remarkable wildlife.
For previous TetZoo articles on birds and birdwatching, see…
To the Sahara in quest of dinosaurs (living and extinct), December 2008
From Morocco, with larks, babblers, gazelles, owls and GIANT DINOSAUR BONES, December 2008
Birding in Brazil: a view from suburban Rio de Janeiro, June 2013
Birdwatching in Suburban China, May 2018
Avocets in Flight and Phylogeny, October 2018
Cocks-of-the-Rock, Extreme Cotingas, April 2019
Refs - -
Chu, P. C. 1994. 1998. A phylogeny of the gulls (Aves: Larinae) inferred from osteological and integumentary characters. Cladistics 14, 1-43.
Cracraft, J. 2014. Avian higher-level relationships and classification: Passeriformes. In Dickinson, E.C. & Christidis, L. (eds) The Howard and Moore Complete Checklist of the Birds of the World (fourth edition), Volume 2: Passerines. Aves Press, Eastbourne, pp. xvii-xlv.
Cracraft, J., Barker, F. K., Braun, M., Harshman, J., Dyke, G. J., Feinstein, J., Stanley, S., Cibois, A., Schikler, P., Beresford, P., García-Moreno, J., Sorenson, M. D., Yuri, T. & Mindell, D. P. 2004. Phylogenetic relationships among modern birds (Neornithes): towards an avian tree of life. In Cracraft, J. and Donoghue, M. (eds) Assembling the Tree of Life. Oxford University Press (Oxford), pp. 468-489.
Selvatti, A. P., Gonzaga, L. P. & Russo, C. A. de M. 2015. A Paleogene origin for crown passerines and the diversification of the Oscines in the New World. Molecular Phylogenetics and Evolution 88, 1-15.
Sibley, C. G. & Ahlquist, J. A. 1990. Phylogeny and Classification of Birds. New Haven: Yale University Press.
Siegel-Causey, D. 1988. Phylogeny of the Phalacrocoracidae. The Condor 90, 885-905.
Hacks Vs Wildlife: the Eternal Vilification of Gulls
Every summer, here in the UK, it’s the same. “CRAZED KILLER SEAGULL ATTACKED MY BABY”, “PSYCHO SEAGULL’S REIGN OF TERROR”, “EXPERT SAYS PUPPIES AND KIDS COULD BE NEXT”. It’s almost as if hack journalists, writing for shitty tabloids like The Mail, The Sun and The Star, have nothing smart to write about so fall back on scaremongering and the vilification of wildlife. Hack journalists are pretty good at this, stirring up waves of anti-gull feelings among a public that already dislikes any animal trying its damnedest to survive and persist in a land dominated by humans and their intolerance of wild places and other species (see #HacksVsWildlife on Twitter for a constant stream of this sort of thing).

Caption: it’s summertime in the UK, and this can only mean one thing….
Right now, the story of Gizmo the chihuahua – apparently snatched by a gull in Paignton, Devon – is doing the rounds. Gizmo is, or was, a very small dog, so the idea that he/she might have been grabbed by a big gull is not out of hand. I’m inclined to think that the event did happen, in which case Gizmo’s owners have my sympathy. Lesson for the future: don’t leave tiny dogs unguarded, outside, in an area where there are big gulls. As others have pointed out, big white-headed gulls like the Great black-backed Larus marinus can swallow prey the size of juvenile rabbits, and a big gull can likely fly while carrying an object about similar in size to a very small chihuahua (cf Young 1987).
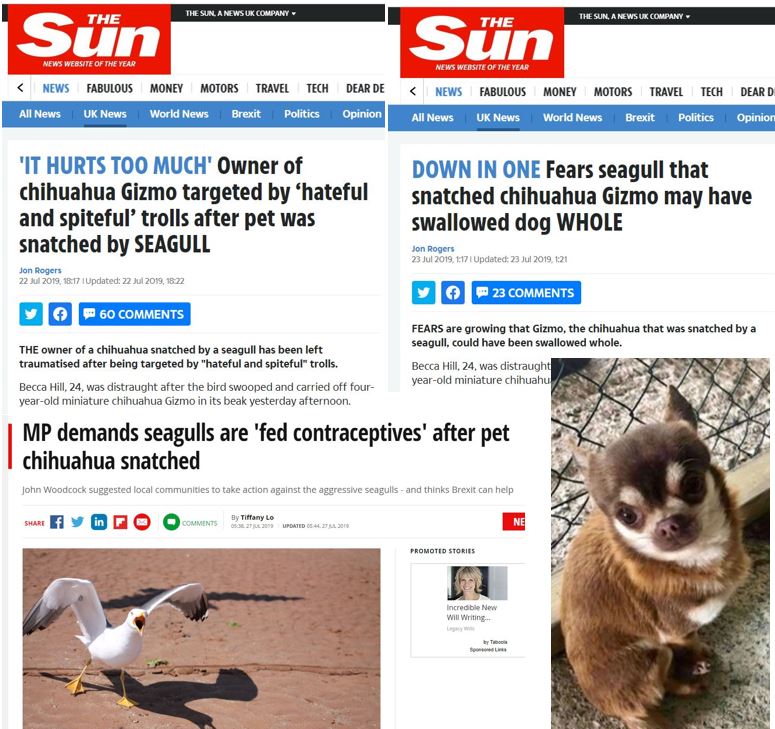
Caption: alas, poor Gizmo. Screengrabs of headlines from some of the UK’s most noble gutter rags.
As cynical as it sounds though, this strikes me as an example of bad pet management more than a case of ‘out of control’ wildlife. I have a pet lizard and guinea-pig who are often taken outside in an area where there are gulls and corvids of several species, and I simply wouldn’t leave them alone and unguarded. What about those parts of the world where there are such things as coyotes, bobcats, eagles and other opportunistic predators? Who is to ‘blame’ when, say, a pet cat is taken by a coyote? Is it really because the coyote is in the ‘wrong’ place?

Caption: white-headed gulls (those gull species remaining in the genus Larus) are good-looking birds, in cases with wingspans that exceed 1.3 m. This is a Herring gull (photographed in Cornwall, England) with an unusually shaped head. Image: Darren Naish.
Are gulls big, formidable and potentially dangerous to small animals and even people? Yes, of course they are. They’re predatory and opportunistic, and often territorial and liable to be aggressive when guarding their nests and chicks. But the idea – promoted continually by hack journalists – that there’s some kind of GULL PLAGUE that we should rid ourselves of is just wrong, and bad.

Caption: Herring gulls consume leftovers at a restaurant in Tintagel, Cornwall. Image: Darren Naish.
Gulls Are In Decline. First of all, let’s look at the idea that these birds are super-abundant, as hack journalists and gull-haters would like you to think. As has now been pointed out many times (here at TetZoo and elsewhere), the species concerned are not just in decline, they’ve declined so much within recent decades that they’re now a cause for conservation concern. In the UK, the Herring gull L. argentatus – the species that hacks and haters mostly focus on – is a Red List species, its population now being at its lowest since recording began in 1969/70, and having declined by about 50% since the early 1990s (Madden et al. 2010, Joint Nature Conservation Committee 2014). The next most familiar species – the Lesser black-backed gull L. fuscus – has undergone a worrying population crash during this century. Similar trends are present in white-headed gulls elsewhere; the North American Herring/Smithsonian gull L. smithsonianus has declined by almost 80% since the 1960s.

Caption: a fine Lesser black-backed gull at a train station. Note the yellow legs, the small extent of the white spotting on the black tips to the primaries, and fairly dark mantle. Image: Darren Naish.
In view of this, it seems wrong to call for these birds to be sterilised (as some politicians have, apparently seriously, suggested), or for them to be lethally controlled or extirpated entirely. I may well be speaking from a position of privilege (I undoubtedly am, in fact, given that I live in a land where there are no big dangerous animals at all), but I want to live in a world where we’re alongside other species, not hell-bent on crushing them under heel into extinction. Urban gulls are an occasional menace for sure, but these aren’t animals that we should vilify or try to expunge. They need help; we should promote tolerance, not destruction.

Caption: a Bristolian Herring gull eating a feral pigeon, again at a train station. Did the gull kill the pigeon, or was the pigeon a victim of a train collision? I don’t know, but either is possible. Image: Darren Naish.
I should also add that urban gulls are important ecosystem service providers, eating waste, carrion and undesirable material (I won’t start listing it, but – oh, ok – it includes vomit and dog scat) left in the built environment by human action. They’re also important seed dispersers, playing this role in areas where other fruit-eating birds (yes, gulls eat fruit) are rare or absent (Iason et al. 1986, Magnusson & Magnusson 2000, Sekercioglu 2006).

Caption: a large group of white-headed gulls compete for food scraps in Lisbon, Portugal. Image: Darren Naish.
Why are people fixated on the idea that we have a ‘gull plague’, or that there are somehow ‘too many’ gulls? I think that a few factors may be at play. One is that gulls are both big and comparatively long-lived, meaning both that they’re way more obvious than small birds, most of which are ignored by the majority of people, and also seen repeatedly in the same areas. A single gull, hanging out on the same area of railway platform or beach promenade for perhaps more than 30 years (a Herring gull ringed in 1965 was still alive in 1997, and older individuals are now on record too), creates the impression of abundance.
There’s A Human Problem, Not A ‘Gull Problem’. Why are gulls so ubiquitous and – to hark back to the hack journalist take – problematic and pestilent? Is it because they’ve devised a clever plan to usurp humans and kill us all by pecking at the face? No, it’s because we’ve created ideal places for them to live, forage and breed thanks to our epic production of useable and edible waste, and our production of (mostly) predator-free, friendly places ideal for resting, feeding and breeding. We’ve also made life harder for them at coasts and seas thanks to development, pollution and industrial fishing. In short, the urban gull ‘problem’ is a direct product of the human problem.

Caption: there’s some degree of uncertainty as goes how reliable urban Herring gull counts are, but the overall trend over recent decades is certainly one of overall population decline. This graph is from the Joint Nature Conservation Committee Herring gull page. The dotted lines show 95% confidence limits. Image: JNCC (original here).
At the risk of repeating myself… yes, big gulls can be less than ideal neighbours. Yes, they’ll steal your chips or sandwiches if you’re unwise enough to hold those things aloft and be unaware of big animals watching you from nearby. Yes, they’ll potentially swoop at you or whack or bite you if you go close to their nests or chicks. Yes, they may even – very, very rarely – do such things as see small dogs and other pets as prey items. Yes, they shit. And, yes, they can be noisy and do their raucous calling at inconvenient times of the day or night.

Caption: white-headed gulls are slow to mature and have different plumage phases depending on age. It’s therefore typical to see birds of several different year stages at any one place where gulls hang out. This 1st winter Herring gull was photographed at Orton, Devon. Image: Darren Naish.
But I seriously question the idea that these often magnificent, slow-growing, long-lived and behaviourally fascinating birds are anything like the ‘problem’ that hack journalists would have us believe. I want to live in a world where there are other animals besides more humans and our domesticates, and the whole idea that gulls are a ‘problem’ is, frankly, tired bullshit that we should be done with.
Gulls and other charadriiform birds have been covered a few times on TetZoo before… though here’s your usual reminder that some of these articles are now paywalled, or have had their images removed.
The godwit’s many bills, January 2007
Why the Lion Grew Its Mane, a book review, November 2008
To the Sahara in quest of dinosaurs (living and extinct), December 2008
Mysterious channels of Alca torda, January 2009
Kleptoparasitism at Westbury Manor, January 2009
Mobile phones, medals, a doll's legs, an entire army... is there anything a gull won't swallow?, December 2009
The incredible bill of the oystercatcher, July 2010
Your Awesome Neighbourhood Herring Gull (And Its Many Cousins), September 2015
Avocets in Flight and Phylogeny, October 2018
Refs - -
Iason, G.R., Duck, C. D. & Clutton-Brock, T. H. 1986. Grazing and reproductive success of red deer – the effect of local enrichment by gull colonies. Journal of Animal Ecology 55, 507-515.
Madden, B. & Newton, S. F. 2010. Herring gull (Larus argentatus). In Lloyd, C., Tasker, M. L. & Partridge, K. (eds) The Status of Seabirds in Britain and Ireland. T & AD Poyser, London, pp. 242-261.
Magnusson, B. & Magnusson, S. H. 2000. Vegetation on Surtsey, Iceland, during 1990–1998 under the influence of breeding gulls. Surtsey Research 11, 9-20.
Sekercioglu, C. H. 2006. Increasing awareness of avian ecological function. Trends in Ecology and Evolution 21, 464-471.
Young, H. G. 1987. Herring gull preying on rabbits. British Birds 80, 630.



