Having recently republished an old, classic article on Speculative Zoology – that based around a 2014 interview with After Man creator Dougal Dixon – it seemed appropriate to republish another Tet Zoo classic…
Speculative Zoology and the World of After Man; an Interview With Dougal Dixon
My Own Personal Dinosauroid
The Snouters or Rhinogradentians, a Brief Review
Humanoid Dinosaurs Revisited Again: Russell and Séguin’s Dinosauroid at (Nearly) 40 Years Old
The Netflix Series Alien Worlds
December 2020 saw the release of the Netflix series Alien Worlds, a four-part exploration of imaginary alien life. The series is science-led throughout and incorporates world-class CG creature effects in addition to real world explorations of the various biological and evolutionary phenomena discussed across the episodes. Sophie Okonedo narrates.
Trope of the Buffalo-Backed Dinosaur
Dougal Dixon’s After Man, the Initial Pitch Document
There can’t be many visitors here who are unaware of Dougal Dixon and his 1981 book After Man (Dixon 1981), the work which effectively started the entire Speculative Zoology (or SpecZoo) movement.
Alternative Timeline Dinosaurs, the View From 2019 (Part 3): the Dinosauroid and its Chums
Welcome to another article on alternative timeline dinosaurs.

Caption: what’s this? It’s an unfinished sculpt of the head of Paranthropoharpax, by Mette Aumala. Read on for more. Image: (c) Mette Aumala.
The previous two articles looked at recent and current ideas on those non-bird dinosaurs that might have evolved in alternative timelines where the end-Cretaceous extinction event never happened, and where the modern world is occupied by the descendants of groups that otherwise went extinct about 66 million years ago. You can see those articles here and here. And the most recent of those two articles looked at the ‘would there be humans anyway?’ argument, always a mainstay of alternative timeline dinosaur discussion.
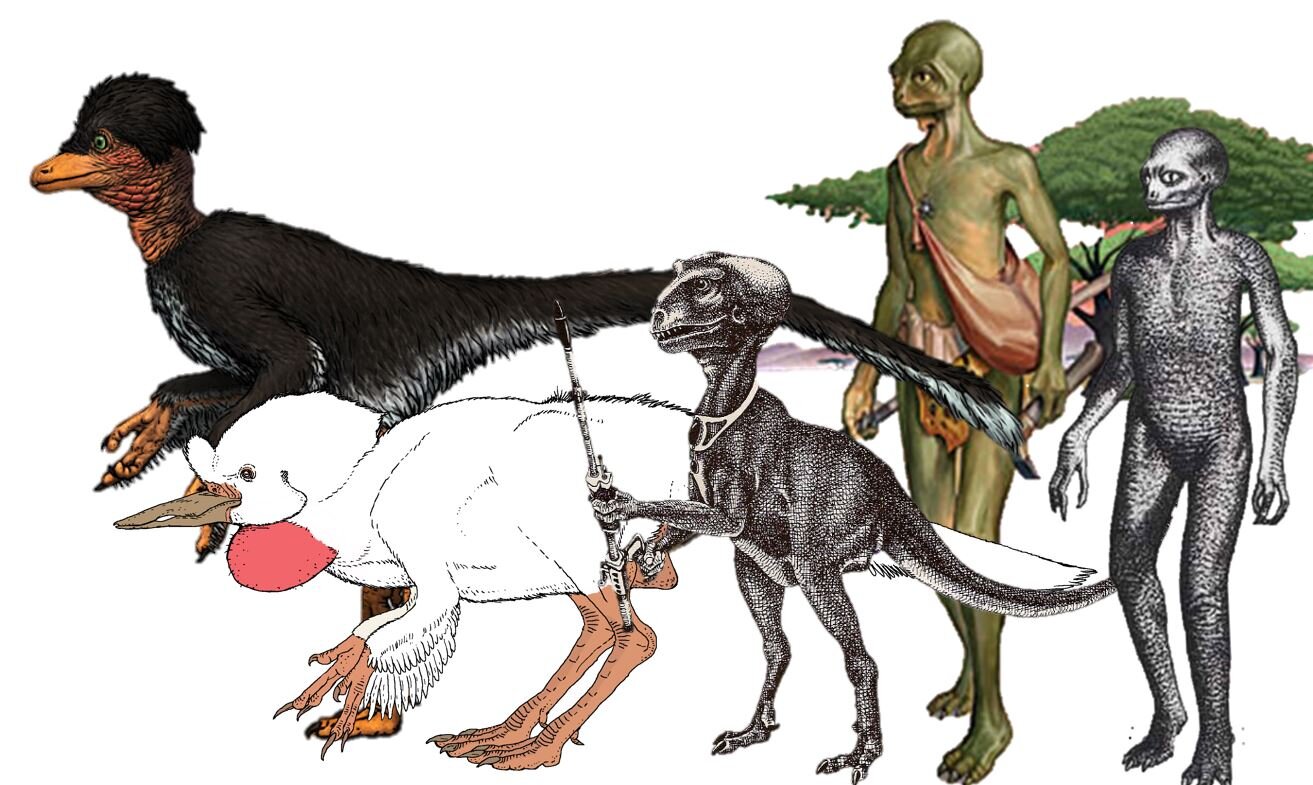
Caption: a whole bunch of dinosauroids, variously by (left to right) Mette Aumala, C. M. Kösemen, John McLoughlin, Matt Collins, John Sibbick.
The other big question that everyone asks about post-Cretaceous, alternative timeline dinosaurs is also about the evolution of intelligence, but this time of a non-human sort. If non-bird dinosaurs didn’t go extinct, surely they’d give rise to animals of primate-like, if not human-like, intelligence… right? The ‘intelligent dinosaur’ meme has been covered quite a bit here on TetZoo before (see links below), the problem as always being that my older articles on this subject are today plagued by hosting issues. For this reason I’ve linked below to the wayback machine versions of the articles concerned.

Caption: if you’ve ever opened a post-1980s dinosaur book, chances are high that you’ve seen photos of Dale Russell and Ron Séguin’s troodontid and dinosauroid models. This is the commonest set of images, as reproduced in Russell (1987, 1989) and other sources.
Conversations about big-brained dinosaurs invariably revolve – as well they should – around Dale Russell’s ‘dinosauroid’ of the 1980s, an imaginary humanoid theropod which Russell posited as a possible evolutionary descendant of troodontid theropods had they continued to evolve beyond the end of the Cretaceous. I’m keen to avoid saying too much about the dinosauroid here for fear of repeating content, but it’s worth noting that Russell’s vision of the dinosauroid was more fleshed out and detailed than many assume, and that its anatomy was more complex and nuanced than might first appear (Russell & Séguin 1982). Anyway, please read the older TetZoo articles if you want to dig deeper.

Caption: a rare photo of Dale Russell (at left) and Ron Séguin in the company of the dinosauroid; this photo belongs to the archives of the Canadian Museum of Nature and was shared in September 2019 by Jordan Mallon. Image: (c) CMN.
The dinosauroid’s infamy was and is due to the construction of an amazingly good life-sized model by sculptor Ron Séguin, who collaborated with Russell on the project (Russell & Séguin 1982). I saw the model in person in 1990 when it came to the UK for the Dinosaurs Past & Present exhibition, but failed to take the illegal photos I now so wish I had. You can read about my recollections of that amazing exhibition here (and be sure to read the comments).

Caption: Ely Kish’s dinosauroid models, used as the basis for a large painting which she finished; it remains in storage and has never seen print. These images were shared on Michael Ryan’s blog (here). Images: (c) Michael Ryan.
Séguin’s construction is well known, but a revelation new(ish) to some of us is that the late Ely Kish – another artist who collaborated with Russell (Russell 1987, 1989) – produced several amazing pieces of art that depict dinosauroids in assorted novel settings. One (a clay sculpture) shows a parent playing with a baby-faced youngster, and another – a large painting – shows a 1980s-era dinosauroid drawing attention to a reconstruction of an artistic scene. That scene is a piece of palaeoart, produced by the dinosauroids, and depicting their own kind during its cave-dwelling, ‘Palaeolithic’ phase of evolution. To my knowledge, the painting has never been published, perhaps because Dale Russell gave up on any dinosauroid-themed projects following the negative response the idea received after appearing in print during the early 80s. It was apparently produced for Russell’s 1989 book An Odyssey in Time: the Dinosaurs of North America (Russell 1989) but eventually excluded.

Caption: some post-Russell versions of the dinosauroid. At far left: a life-sized model on display at Dorchester’s Dinosaur Museum; at centre, two views of the suit made by Peter Minister for the 1991 TV series Dinosaur!; at far right, John Sibbick’s take on the dinosauroid, from David Norman’s 1985 book The Illustrated Encyclopedia of Dinosaurs. Images: Jim Limwood, CC BY 2.0 (original here), Norman (1991), John Sibbick/Norman (1985).
The dinosauroid has proved such a mainstay of post-1980s dinosaur pop-culture that it’s mentioned and discussed in a vast number of popular books on dinosaurs and prehistoric life, and has often been redrawn (often incorrectly); models and even suits worn by actors have been constructed too. One day I should compile a bibliography. And dinosauroid-like animals have been invented by several writers and authors beyond Dale Russell, among them John McLoughlin’s big-brained dromaeosaur (McLoughlin 1984), Magee’s Anthroposaurus (Magee 1993), the hadrosaurian Voth of Star Trek: Voyager, and Mette Aumala’s Paranthropoharpax naishi.

Caption: at left, John McLoughlin’s dinosauroid (which I opted to name Bioparaptor). At right: the cover of Magee’s unusual book on big-brained dinosaurs (which Amazon now values at about £200). Images: McLoughlin 1984, Magee 1993.
I still say that the dinosauroid sucks. Its entire look is dependent on the idea that the human shape is the ‘best’ one for an intelligent, big-brained animal, an idea that Russell appears to find palatable if not preferable (Russell 1987, p. 130; Psihoyos & Knoebber 1994, p. 252). But this doesn’t mean that the entire concept of big-brained smart dinosaurs is a bad one. As has now been said many times, birds (parrots and corvids, at least) overlap with primates in relative brain size and cognitive abilities, meaning not only that big-brained smart dinosaurs evolved for real in our own timeline, but also that it could happen again in a timeline where other bird-like lineages exist.
And if big-brained, smart non-bird theropods really evolved, they would – I think – more likely be shaped like birds and other theropods: not erect-bodied and humanoid, but horizontal-bodied, feathery, and with the long-snouted faces and inward-facing, clawed hands more typical of the group. This argument is old hat now since it got a lot of coverage back in 2006 when C. M. Kösemen (aka Memo) – inspired, I think, by my writings on alternative timeline smart dinosaurs – invented Avisapiens saurotheos.
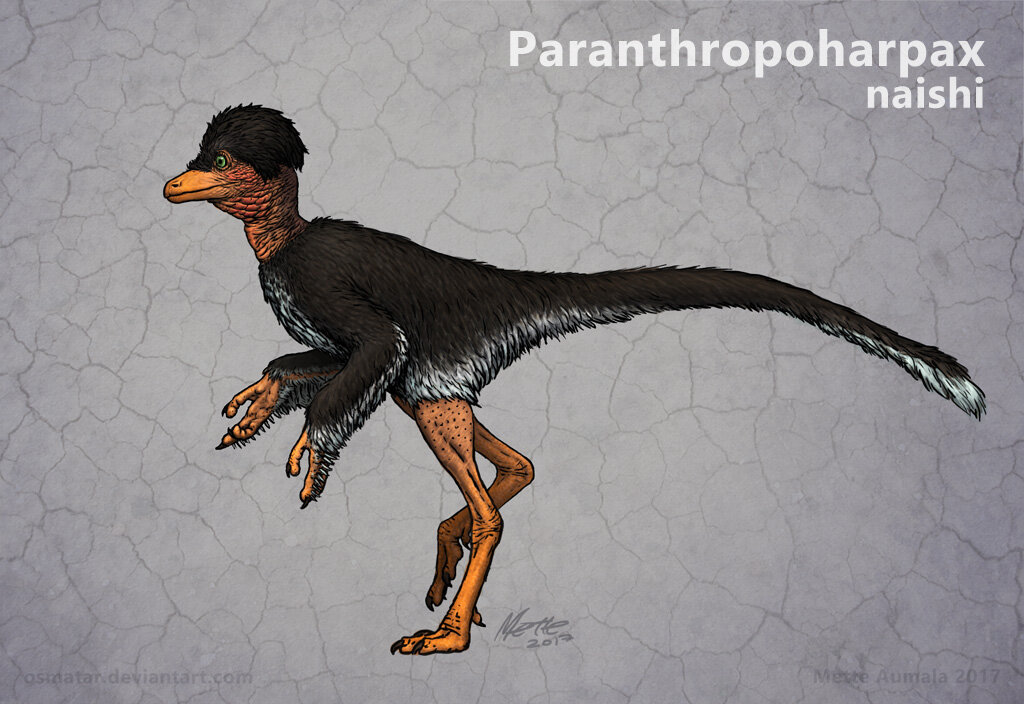
Caption: for a change, I’m not going to share Memo’s Avisapiens (well, it’s already visible in the montage at top), but Mette Aumala’s Paranthropoharpax naishi. For an article on this creature go here. Mette has recently been working on a CG portraits of this species (go here on Twitter). Image (c) Mette Aumala.
In the years since 2006, Memo has invented additional big-brained hypothetical dinosaurs and – via collaboration with Simon Roy – has devised a parallel world where big-brained dinosaurs of more than one species possess their own culture, technology and art, and are depicted alongside beasts of burden and other contemporaries. Post-2006, we’ve also seen the coming and going of the 2007 kid’s TV series Dinosapien, the appearance of some surprisingly pro-dinosauroid commentary from Richard Dawkins (I wrote about that event here), and the coverage of the dinosauroid in numerous articles and books (e.g., Hecht 2007, Naish 2008, Socha 2008, Switek 2010, Losos 2017).

Caption: screengrabs of pages from two recent-ish articles discussing the dinosauroid: Hecht (2008) and Socha (2008).
And the big-brained dinosaur trope continues to raise its head in the popular and semi-technical sphere, often from people who seemingly aren’t aware of Russell’s dinosauroid and appear to be discovering it for the first time. The increasing popularity of ideas about humanoid reptilian aliens and a cryptic elite of shape-shifting lizard-people (good work, David Icke) mean that people whose idea of research starts and ends with Google are discovering dinosauroids and somehow working them into a world view that involves the Illuminati and high-level One World Government conspiracies. In short, it’s an idea that isn’t going away, and Jonathan Losos was right in saying that “… to my surprise … The dinosauroid hypothesis was alive and thriving in cyberspace” (Losos 2017, p. 323).
And that, effectively, brings us up to date…. for now. What might happen next in the world of Alternative Timeline Dinosaurs?
For previous TetZoo articles on alternative timeline dinosaurs and related issues of SpecZoo, see…
Dinosauroids revisited, November 2006
Dinosauroid cave art discovered, March 2007
How intelligent dinosaurs conquered the world, March 2008
Come back Lank, (nearly) all is forgiven, September 2008
Belatedly, Nemoramjetia (= Avisapiens), November 2008
Richard Dawkins and the crappy 'humanoid dinosaurs' that just won't die, November 2009
Dinosauroids revisited, revisited, October 2012
Of After Man, The New Dinosaurs and Greenworld: an interview with Dougal Dixon, April 2014
Speculative Zoology, a Discussion, June 2018
Could We Domesticate (Non-Bird) Dinosaurs?, August 2018
The Dougal Dixon After Man Event of September 2018, September 2018
TetZoo Bookshelf, February 2019, Part 1, February 2019
Alternative Timeline Dinosaurs, the View From 2019 (Part 1), December 2019
Alternative Timeline Dinosaurs, the View From 2019 (Part 2), December 2019
Refs - -
Hecht, J. 2007. Smartasaurus. Cosmos 15, 40-41.
McLoughlin, J. 1984. Evolutionary bioparanoia. Animal Kingdom April/May 1984, 24-30.
Naish, D. 2008. Intelligent dinosaurs. Fortean Times 239, 52-53.
Norman, D. 1991. Dinosaur! Boxtree, London.
Psihoyos, L. & Knoebber, J. 1994. Hunting Dinosaurs. Cassell, London.
Russell, D.A. & Séguin, R. 1982. Reconstruction of the small Cretaceous theropod Stenonychosaurus inequalis and a hypothetical dinosauroid. Syllogeus 37, 1-43.
Socha, V. 2008. Dinosauři: hlupáci, nebo géniové? Svĕt 3/2008, 14-16.
Alternative Timeline Dinosaurs, the View From 2019 (Part 2)
In the previous article we looked at alternative timeline dinosaurs, and in particular at Dougal Dixon’s The New Dinosaurs, at the Speculative Dinosaur project, and at a few things that connect these two endeavours.

We finished that article by introducing an idea mentioned or discussed several times in discussions about SpecZoo: given that mammal evolution would surely continue in a dinosaur-dominated, post-Cretaceous world, could big-bodied mammals evolve, and could they give rise to humans... or, at least, to human-like primates?
Before I continue, I should add that this issue is especially topical at the time of writing (early December 2019) since a BBC World Service radio programme featuring this very issue has just appeared. It’s part of the CrowdScience series, is titled ‘Would humans exist if dinosaurs were still alive?’ and can be found here. It features me, Memo Kosemen, Elsa Panciroli, Anjali Goswami and Nicola Clayton.

Caption: a whole radio show on speculative dinosaurs! You need a BBC account to access it. Image: BBC (from here).
Could there be humans? Well, this would require the (mostly) terrestrial evolution of a big-bodied primate group, and I said in the previous article (where I called it the no megamammals rule) that that might not be possible in a dinosaur-dominated world (and I mean because dinosaurs and other archosaur groups were filling up the available niche space, not because predatory species were perpetual eliminators of animals that dared set foot on the ground or anything like that). Could, then, we have hypothetical ‘human-level’ apes (thinking here of cultural and technological sophistication, and/or relative intelligence) evolve in an arboreal setting?
Here it’s worth saying that the existence of hypothetical climbing and flying predatory archosaurs might not be an evolutionary obstacle to the existence of relatively big arboreal mammals. I say this because large, scansorial and arboreal mammals have already evolved in a world where they’re predated upon by large, predatory flying theropods, some of which can reach into cavities with flexible legs and feet, or pull relatively big prey items (like sloths and big monkeys) from treetops and branches.
But… if I let my archosaur bias run away with me, I might propose that the existence of arboreal mammals in a dinosaur-dominated world could encourage the evolution of specialised scansorial or arboreal archosaurs (presumably theropods) that have somehow influenced primate evolution, and ultimately derailed any potential evolution of proto-humans. I like this idea, and it isn’t entirely without precedent in our own timeline, since it’s been formally suggested that the predation pressure exerted upon primates by large predatory reptiles – snakes, specifically – has indeed shaped primate evolution (Isbell 2006).

Caption: a speculative arboreal maniraptoran from an alternative timeline. It runs and leaps about in tropical tree-tops, has extremely powerful, prehensile feet, and predates on primate-like mammals. Image: Darren Naish.
Here’s another possibility: could human-like apes evolve on an island refugium where archosaurs weren’t dominant? The idea that ‘unusual’ animals can evolve and persist on islands is already a mainstay of island biogeography theory and verified by many real-world examples. A dinosaur-dominated post-Cretaceous world must be allowed to include similar diversity, and maybe this is where you find your big, atypical mammals (and perhaps big tortoises, arthropods and whatever else).
Could human-like primates evolve on islands? Well, this would require that your ancestral primates occurred in such places in the first place, and that the relevant selection pressures would have led to the evolution of human-like animals. While this isn’t impossible, it seems unlikely: there are good reasons for thinking that the ecological conditions specific to certain regions of Neogene continental Africa are what ultimately resulted in the emergence of hominins and hence humans (Kingdon 2003). My conclusion here is that an alternative evolutionary pathway for primates doesn’t give you humans as one of its ‘products’.
Finally, if we have an alternative timeline where apes evolve, is it conceivable that human-like primates, and, ultimately, alternative timeline humans, evolve anyway, dinosaurs or not? After all, humans and human ancestors spent their entire history living alongside formidable predators and competitors which either had to be avoided, defeated or out-smarted, and we might argue that the technological and other behavioural sophistications of hominids could make them exempt from the ‘no megamammal rule’ introduced above. The result would be what I’m going to call a Flintstones Universe.

Caption: non-bird dinosaurs and humans live alongside one another in the episode In Dino Veritas (season 2, ep 7) of the TV series Sliders, which aired in April 1996.
As appealing as this model might be for the purposes of human-oriented drama and adventure (it’s been done before, in Sliders - a TV series about travel between alternative timeline versions of Earth - and I’m sure elsewhere), I’m worried, since it also sounds a lot like the ‘inevitable humans’ model promoted by those who seem to regard humans as The Best Animals. Some of you might have seen the 2010 episode of the BBC TV series Horizon (it’s here) where Simon Conway Morris – who’s well known, these days, as something of an opponent to the late Stephen Jay Gould’s argument that nothing in evolution is inevitable and that events wouldn’t have run the way they did if we were to “replay life’s tape”* (Gould 1990, p. 48) – proposed both that humans could well have evolved in a dinosaur-dominated world and that smart humanoid dinosaurs could well have evolved as well. Even better, Conway Morris suggests in the same TV show that humans and big-brained dinosaurs could have forged an alliance. True, true, symbiosis is not uncommon in the living world – ants and aphids, cleaner fishes and big reef fishes, Godzilla and Mothra – but the real-world history of humans shows either that we smashed competitors into bloody fragments or absorbed them into our collective via sexy, sexy ways. The latter option is unlikely to play out in a world where humans and humanoid dinosaurs live together, but the former suggests a less than happy outcome for one of the players.

Caption: some of the key books relevant to the discussion here.
So, excuse me if I’m a little sceptical of any proposed human-dinosauroid buddy-flick partnership. And am I sceptical of the idea that hominids, hominines and hominins were indeed exempt of the ‘no megamammals rule’? Yes. Incidentally, Conway Morris’s inevitability vs Gould’s contingency formed the subject of a long-running dispute: Riley Black wrote about this discussion here.
* Does this analogy still work given that hardly anyone uses recording tape anymore?

Caption: Simon Conway Morris talks about the evolution of intelligence while a dinosauroid reads a newspaper in the background. This is a screengrab from the 2007 episode of BBC Horizon titled My Pet Dinosaur.
Other, more recent alternative timeline dinosaur thoughts. As should be clear from this article and its predecessor, interest in alternative timeline dinosaurs is at an all-time high. The current resurgence of monster movies that feature King Kong, Godzilla and the like also means that such creatures have appeared in big-budget, mainstream movies. Peter Jackson’s 2005 movie King Kong featured several post-Cretaceous dinosaurs (among them Vastatosaurus rex, Brontosaurus baxteri and Venatosaurus saevidicus), the design and backstory of which makes at least some sense on the basis of things that happened in the real timeline… though the idea of a dinosaur genus persisting relatively unchanged for 150 million years or so can’t be considered sensible in view of what we know.

Caption: I don’t especially like the dinosaurs of Jackson’s King Kong (they’re too conservative), but a lot of the artwork is neat. This concept art depicts Vastatosaurus rex, a giant Holocene tyrannosaurid that evolved from Tyrannosaurus. Image: The World of Kong (here).
Pixar’s 2015 The Good Dinosaur also has to be mentioned, but I wish it didn’t because I think it’s a terrible film and an abhorrent waste of an opportunity. It’s supposed to be an alternative timeline movie but instead it just seems like a Flintstones reboot without the Flintstones.

Caption: opening spread from Pickrell (2017), art by James Gilleard. Some of the creatures here are based on those of TND and the Speculative Dinosaur Project. Image: BBC Focus magazine.
A 2017 article by John Pickrell is one of the newest published pieces on alternative timeline dinosaurs, and it’s well worth tracking down for those seriously interested in this stuff (Pickrell 2017: there’s an online version here which includes some extra stuff relative to the printed one). It features some fairly outré artwork (by James Gilleard; it’s not included in the online version) depicting creatures inspired by those of The New Dinosaurs, and include quotes from Steve Brusatte, Tom Holtz, Andrew Farke, Matthew Bonnan and Victoria Arbour (and Paul Barrett and me, but in the online version only) (Pickrell 2017).

Caption: the Gourmand Ganeosaurus tardus, a carrion-feeding tyrannosaur whose ancestors migrated to South America during an alt-timeline version of the Great American Biotic Interchange. I’ve never been able to figure out what it’s eating. Image: art by Steve Holden, from Dixon (1988).
A few interesting, specific speculations are made in John’s article. Tom notes that the Gourmand of TND is maybe not so ridiculous given recent discoveries concerning abelisaurs (rather than tyrannosaurs), Matthew suggests that arboreal (non-bird) dinosaurs might have co-evolved with flowering plants, and Victoria proposes that the neornithine bird radiation might not have been as bushy and explosive if pterosaurs had never died out (Pickrell 2017). Other ideas mooted in the article are that climbing, monkey-like theropods might evolve, and that there might be shaggy-coated Arctic specialists, speedy grassland herbivores and whale-like descendants of spinosaurs; in part, they’re seemingly inspired by the Dixonian creatures of TND.

Caption: more art from Pickrell (2017), by James Gilleard. Image: BBC Focus magazine.
The online version of the article includes comments on some of the events of post-Cretaceous times that – if they happened in an alternative timeline – would likely have some impact on the evolution of dinosaurs, pterosaurs and so on. Examples: what would the Paleocene-Eocene thermal maximum mean for dinosaurs, how might dinosaurs and pterosaurs respond to the cooling and drying of the Miocene, the spread of grasslands and so on, and what might happen in the cool, dry conditions of the Pleistocene? There’s vast scope for speculation here and even for some actual science, since we have enough data to show that organisms evolve in predictable directions when faced with increasing or decreasing temperatures and other such changes.
It's at this point that I have to stop – but there’s more to come. The next article (last in this series) covers ‘dinosauroids’ and other hypothetical big-brained dinosaurs.
For previous TetZoo articles on alternative timeline dinosaurs and related issues of SpecZoo, see…
Dinosauroids revisited, November 2006
Dinosauroid cave art discovered, March 2007
How intelligent dinosaurs conquered the world, March 2008
Come back Lank, (nearly) all is forgiven, September 2008
Belatedly, Nemoramjetia (= Avisapiens), November 2008
Richard Dawkins and the crappy 'humanoid dinosaurs' that just won't die, November 2009
Dinosauroids revisited, revisited, October 2012
Of After Man, The New Dinosaurs and Greenworld: an interview with Dougal Dixon, April 2014
Speculative Zoology, a Discussion, June 2018
Could We Domesticate (Non-Bird) Dinosaurs?, August 2018
The Dougal Dixon After Man Event of September 2018, September 2018
TetZoo Bookshelf, February 2019, Part 1, February 2019
Alternative Timeline Dinosaurs, the View From 2019 (Part 1), December 2019
Refs - -
Dixon, D. 1988. The New Dinosaurs: An Alternative Evolution. Salem House Publishers, Topsfield, MA.
Isbell, L. A. 2006. Snakes as agents of evolutionary change in primate brain. Journal of Human Evolution 51, 1-35.
Pickrell, J. 2017. What if the dinosaurs had survived? In Bennett, D. (ed) Dinosaurs: the Ultimate Guide to the Prehistoric Beasts. Immediate Media, Bristol, pp. 74-83.
Alternative Timeline Dinosaurs, the View From 2019 (Part 1)
Speculative Biology, Speculation Zoology and Speculative Evolution (technically speaking, they’re not all the same one thing) have been perennially popular here at TetZoo, and I think it’s fair to say that interest in these issues is currently at an all-time high. And among the various branches of the SpecVerse that enjoy the most attention and interest is that concerning Alternative Timeline Dinosaurs… and pterosaurs, and marine reptiles, and their contemporaries. These being those hypothetical animal lineages that might have evolved in a timeline where the end-Cretaceous extinction event (properly the KPg Event) didn’t occur.
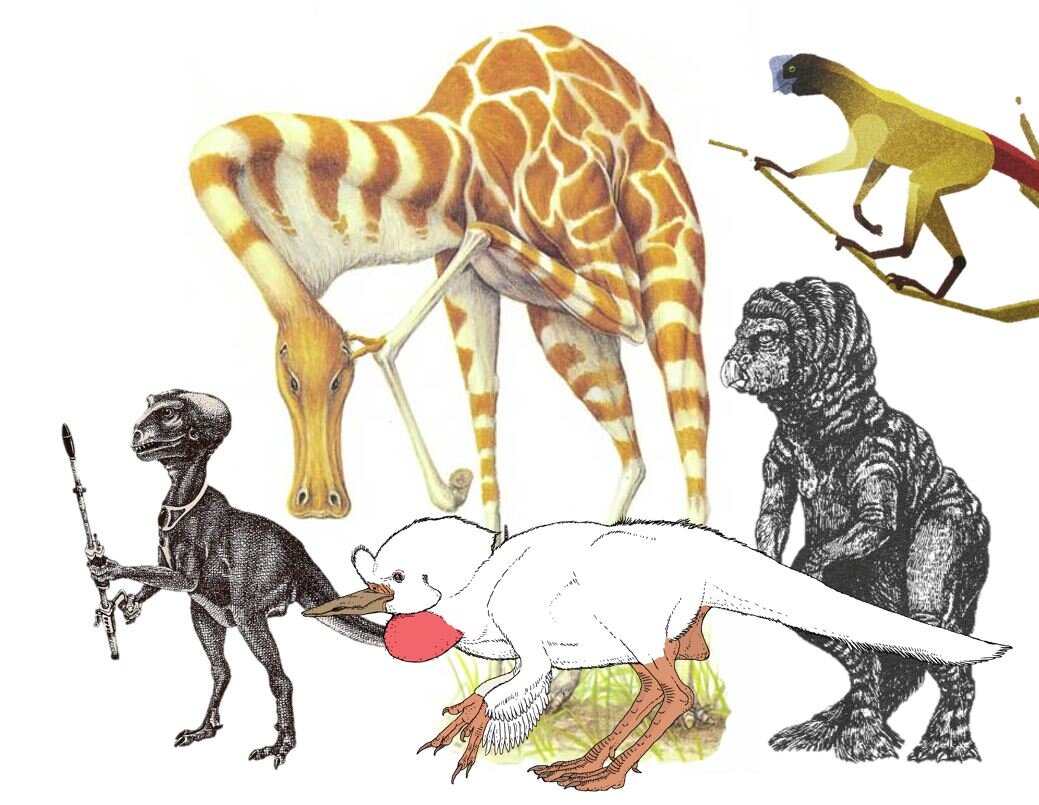
Caption: a montage of speculative dinosaurs, and one pterosaur. From left to right: McLoughlin’s Bioparaptor, Dixon’s Lank (at back), Kosemen’s Avisapiens, Dixon’s Balaclav, and a tree maniraptoran from Pickrell’s 2019 article. Some of these animals will be discussed in future TetZoo articles.
Such is the popularity of alternative timeline dinosaurs that new articles, opinions, ideas and artworks devoted to them are published on a fairly regular basis. At least some of these projects tend not – in my opinion – to give appropriate credit or mention to what’s happened before, but others certainly do. My aim in this article here (and its successors, to be published later this year) is to discuss most of the key ideas that have been covered in takes on alternative timeline dinosaurs, and to review and cite relevant literature old and new. To work.

Caption: the benchmark in the world of alternative timeline dinosaurs is this book, appearing in 1988… the same year that one or two other influential books on dinosaurs appeared in print (looking at you, Predatory Dinosaurs of the World). Image: Darren Naish.
It’s fair to say that hypothetical post-Cretaceous non-bird dinosaurs have featured in fictional scenarios for almost as long as non-bird dinosaurs have been known to science, such that they star in the works of Arthur Conan Doyle, Edgar Rice Burroughs and others since well prior to the dinosaur renaissance. As significant as these works are, however, they mostly don’t get classed as SpecZoo given that they aren’t contingent on the idea that the animals have changed since Mesozoic times. With that in mind, we mostly owe the genesis of the alternative timeline dinosaur trope to two pieces of literature: Dougal Dixon’s The New Dinosaurs: An Alternative Evolution of 1988, and Dale Russell and Ron Séguin’s 1982 Syllogeus article on hypothetical big-brained theropods.

Caption: my introduction to alternative timeline speculative dinosaurs was in part thanks to David Lambert’s Dinosaur Data Book (Lambert 1990), which features this really nice montage: there are Dixonian dinosaurs from The New Dinosaurs, and Russell and Séguin’s dinosauroid at far right. The brief BBC Wildlife magazine ad on The New Dinosaurs - featuring the Kloon (top right) - also caught my imagination. For all kinds of reasons, I never saw The New Dinosaurs in bookshops. Images: Dixon 1988, Lambert 1990.
Dixon (1988) – the sequel to Dougal’s hugely successful and highly popular After Man of 1981 – portrays a modern world where dinosaurs, pterosaurs, plesiosaurs and other groups occupy the roles taken today by placental and marsupial mammals, as well as by various snakes, birds and so on. I won’t start reviewing the pantheon of creatures covered in the book here, but the illustrations below gives some idea of what we get.

Caption: a montage that depicts various of the creatures of Dougal Dixon’s TND. We see here a combination of flightless pterosaurs, climbing and terrestrial, predatory theropods, aquatic ornithischians, amphibious pterosaurs and theropods, and others. These illustrations are redrawn from those featured in Dixon (1988). Image: Darren Naish, colouring by Ethan Kocak.
I think it’s fair to say that The New Dinosaurs (TND) enjoyed a mixed response, with some reviewers hating it and others praising it (e.g., Gee 1988, Tudge 1988, Unwin 1992). The most interesting and detailed review (and, for a long time, the most difficult to obtain) is that by Greg Paul (1990). Paul was highly critical (perhaps ironically, given that his own writings and illustrations were partly inspirational) and made the interesting point that some of the most embracing of Dougal Dixon’s colleagues – Desmond Morris was one of them (he wrote the foreword) – were neither palaeontologists nor especially connected to studies of Mesozoic life. A bonus feature of Paul’s review is that it includes a novel illustration depicting his own take on alternative timeline dinosaurs, his ideas (Paul 1990) being interesting and innovative enough that they’ve inspired more recent views, on which read on.

Caption: alternative timeline dinosaurs of the modern era, as imagined by Greg Paul (1990). Plains-dwelling hadrosaurs and ceratopsians are hunted by cursorial tyrannosaurs. They live alongside small, cud-chewing bipedal ornithischians, burrowing mammals and geese. Image: (c) Greg Paul.
But enough on the critical reception. If anything, the pendulum has swung the other way in recent years, it being increasingly recognised that at least some of the creatures invented for TND were actually insightful and predictive. The Lank – a flightless, giraffe-like, striding pterosaur adapted to life on the grasslands – now appears oddly prescient in view of our thoughts on the ecology and lifestyle of the long-limbed, terrestrially competent azhdarchids (Witton & Naish 2008), and I wrote a 2008 article making this point. The well insulated, cool climate maniraptoran theropods and chubby, insulated ornithischians of TND also no longer look as weird as they might have in 1988 given recent fossil finds, and real-world discoveries like alvarezsaurids, scansoriopterygids, Halszkaraptor and Liaoningosaurus certainly have a Dixonian vibe to them. This was Riley Black’s point, made in this 2011 article at Laelaps.
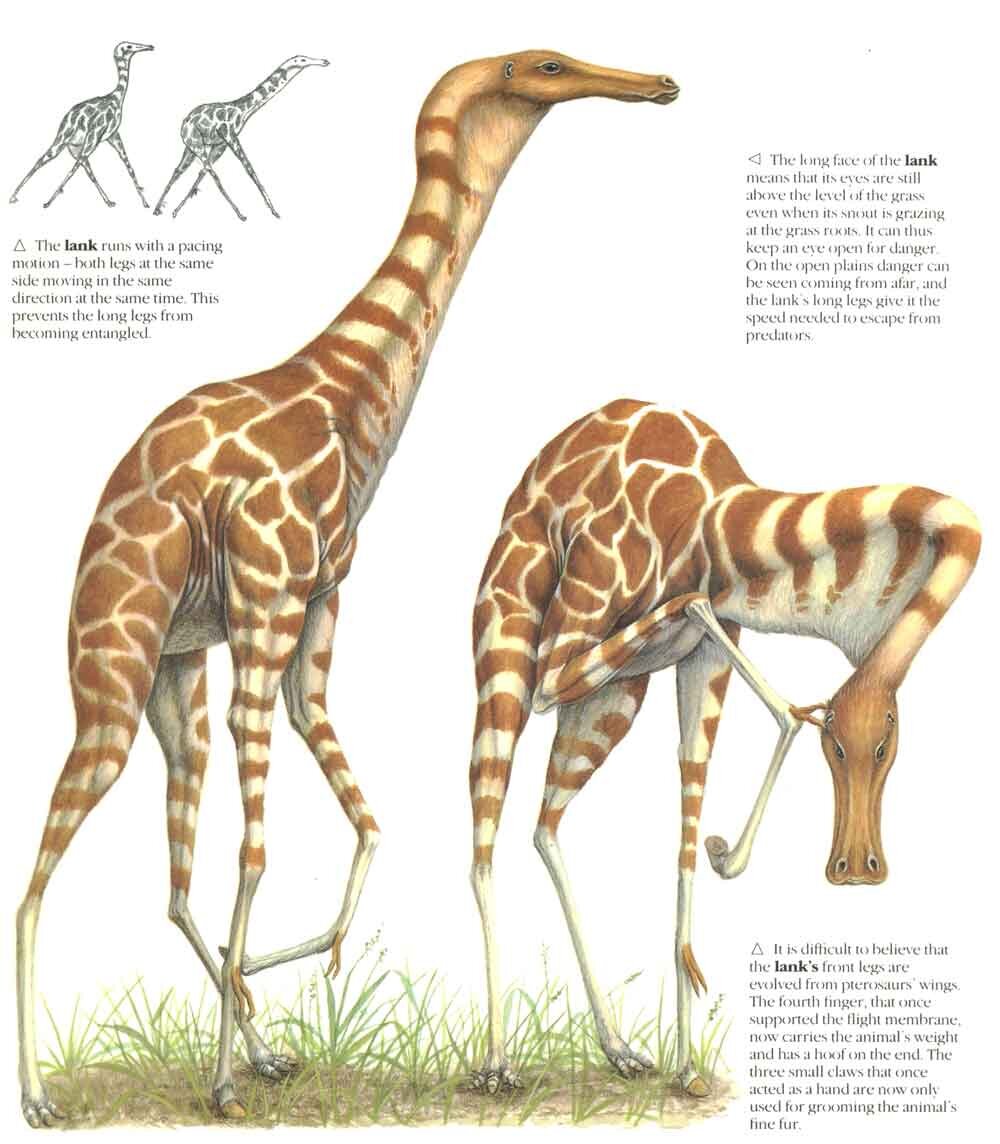
Caption: the Lank has sometimes been criticised as one of the worst creatures of TND. But maybe this isn’t at all fair… Image: (c) Dixon 1988/Steve Holden.
Of Manga and SpecWorld. There are two other things worth saying about TND before we move on. One is that several of the species in the book appeared in a 2006 Manga volume, illustrated by Takaaki Ogawa (Dixon & Ogawa 2009). I tweeted about the book here; it’s hard to get but is a must-have if you’re a serious SpecZoo aficionado.

Caption: at left, the cover of Dixon & Ogawa (2009). At right, one of the several colour pieces included within. Images: (c) Dixon & Ogawa (2009).
The stories in the book are great fun (and you don’t need to read Japanese to follow them), but often very odd because they show the animals of the book living alongside, and interacting with, real-world animals, including crows, arapaima and Passenger pigeons Ectopistes migratorius! That’s a no-no if we’re keeping to canon, since TND actually describes a world where ‘Mesozoic’ animals have evolved to fill the niches occupied in our timeline by other animals. The second thing worth saying is that Dougal’s visions of alternative timeline dinosaurs inspired others to do the same, but in different fashion.

Caption: scenes from Dixon & Ogawa (2009), one from a Harridan-themed story (the Harridan is a raptor-like, mountain-dwelling pterosaur), and one from an Amazon-themed story involving a Pangaloon and a Watergulp. Images: (c) Dixon & Ogawa (2009).
The Speculative Dinosaur Project kicked off in 2001 and involved the invention of a substantial number of speculative animals. The website is impressive and well designed, and it’s clear that a lot of thought and discussion went into the various animals. They include long-legged, often shaggy-coated tyrannosaurs, the artiodactyl-esque ungulapedes, the sometimes gigantic cenoceratopsians (which, like Greg Paul’s alt-timeline ceratopsians, descend from non-ceratopsid ceratopsians) and many others. There’s always been talk of a Speculative Dinosaur Project book, interest in the site remains high, and there are indications right now from various minor tweaks and updates that it might be undergoing an upgrade or something sometime soon.

Caption: brilliant artwork appears throughout the Speculative Dinosaur Project. This illustration depicts a group of Eastern balundaurs, with mulongs, perfects and jaubs in attendance. Image: CC BY-SA 2.5, Speculative Dinosaur Project (original here).
A nice thing about the Speculative Dinosaur Project is that it never was devoted to non-bird dinosaurs alone, but also covered birds, crocodylomorphs, squamates and mammals in appropriate detail too. In fact, the pages on those groups were among the best parts of the site. This leads us nicely to the next area I want to discuss.
Don’t Ignore the Mammals. Of the several questions always asked in discussions about alternative timeline dinosaurs, one is ‘would humans have evolved, even in a world still dominated by non-bird dinosaurs?’. This is a good question, even if its motivation is that we might get to imagine medieval knights using dinosaurs as mounts, a line of armour-plated war ceratopsians at the Battle of Ipsus, or wonder about the role of giant dinosaurs as beasts of burden or quarry hunted by intrepid human bands equipped with specialised weapons.
The human/dinosaur coexistence thing formed the focus of a 2019 Italian magazine article by Giovanni Camardo (Camardo 2019) – titled ‘If they hadn’t died out, who would have won: us or them?’ – for which I was interviewed (the full text of my interview – in Italian – is here).

Caption: opening pages of Camardo (2019). By now, James Keuther’s CG dinosaurs might be somewhat familiar (I’ve worked with James on several projects, most notably various recent Dorling Kindersley books). UPDATE: thanks to Caleb W. (see comments), I now know that the the model was created by Raul Lunia, but then used by others. Image: (c) Focus magazine.
My suggestion – which was inspired by the Greg Paul article discussed above – is that the Cretaceous record of fossil mammals shows us both that interesting things were happening in mammal evolution late in the Mesozoic, and that at least some of the major mammal lineages were already in existence before the end of the Cretaceous. Ergo, at least some of the things that happened for real in our own timeline might still have happened in a dinosaur-dominated post-Cretaceous world. Because non-bird dinosaurs would have occupied most terrestrial niches in post-Cretaceous world, it might – I suggest, with caveats – have been difficult or impossible for mammals to get an evolutionary toehold at large body size (say, above 30-40kg or so). I’m going to call this the no megamammals rule (we’ll come back to it in the next article).

Caption: the Speculative Dinosaur Project was and is particularly good on mammals. Here are two I chose at random: the Amazonian Nekopossum (left) and the apocryphal Drop-bear, also native to South America (unlike our own timeline’s Drop-bear). Image: Speculative Dinosaur Project.
Lineages producing species below this size, however, could still have evolved pretty much as they did in our own timeline. Which means that burrowing, swimming, climbing, gliding and flying mammals – monotremes, placentals and marsupials – might still have evolved in a dinosaur-dominated world, and that a post-Cretaceous world dominated by dinosaurs might still have pangolins, shrews, moles, bats…. and primates. And if there are primates, there might still be tarsiers, lemurs, monkeys and even apes. And if there are apes, could there be the apes we call humans?
This is where we’ll pick up next. Come back soon for Part II.
For previous TetZoo articles on alternative timeline dinosaurs and related issues of SpecZoo, see…
Dinosauroids revisited, November 2006
Dinosauroid cave art discovered, March 2007
How intelligent dinosaurs conquered the world, March 2008
Come back Lank, (nearly) all is forgiven, September 2008
Belatedly, Nemoramjetia (= Avisapiens), November 2008
Richard Dawkins and the crappy 'humanoid dinosaurs' that just won't die, November 2009
Dinosauroids revisited, revisited, October 2012
Of After Man, The New Dinosaurs and Greenworld: an interview with Dougal Dixon, April 2014
Speculative Zoology, a Discussion, June 2018
Could We Domesticate (Non-Bird) Dinosaurs?, August 2018
The Dougal Dixon After Man Event of September 2018, September 2018
TetZoo Bookshelf, February 2019, Part 1, February 2019
Refs - -
Camardo, G. 2019. Se non si fossero estinti, avremmo vinto noi o loro? Focus 317, 51-55.
Dixon, D. 1988. The New Dinosaurs: An Alternative Evolution. Salem House Publishers, Topsfield, MA.
Dixon, D. & Ogawa, T. 2009. The New Dinosaurs. Futanasha, Tokyo.
Gee, H. 1988. Tales of future past. Nature 335, 505-506.
Lambert, D. 1990. Dinosaur Data Book. Facts on File, New York.
Paul, G. S. 1990. An improbable view of Tertiary dinosaurs. Evolutionary Theory 9, 309-315.
Tudge, C. 1988. End points of an alternative evolution. New Scientist 120 (1641), 65-66.
Unwin, D. M. 1992. The New Dinosaurs: An Alternative Evolution (review). Historical Biology 6, 61-71.
Godzilla: King of the Monsters, Some Trivial But Monstrous Thoughts
I’m a big fan of Godzilla, and of kaiju movies in general. So it shouldn’t be a surprise that I just went and saw Godzilla: King of the Monsters (KOTM), and – boy – did I like it. I don’t care about criticisms concerning the human plot (which I thought was fine, perfectly serviceable and well-acted), nor do I care that various possible plot-holes perhaps create issues. Look, it’s a film about giant monsters - rebranded titans for KOTM - knocking the crap out of each other, stomping on cities, out-flying military jets and messing about with lava, ice, epic weather, electricity and nuclear power. I loved it, and I applaud the bravery of the team concerned in taking classic monsters like King Ghidorah and remaining mostly faithful to their design without the need for some major overhaul.

A main theme of Godzilla: King of the Monsters is that humanity is an infection upon the planet, and that titans are the cure. Image: (c) Warner Bros.
I noticed a lot of interesting trivial little details in the film that are of interest or relevance to readers of this blog, at least some of which will have been missed unless you’re a monster fan or have serious nerdy interests in zoology. I mean, there are a hundred other things that are references to other stuff relevant to the Godzilla films and their real-world, factual backstory (Serizawa’s watch, the reference to a Steve Martin, Castle Bravo, Infant Island, maser turrets, the term ‘Monster Zero’, the use of a frikkin’ oxygen destroyer… and so on). It’s the more zoology-themed things I mostly want to highlight here.
The ‘science of Godzilla’ has been covered a fair bit at TetZoo in the past… Image: The Biological Nature of Godzilla.
I should add that Godzilla has been covered at TetZoo on several previous occasions, the usual caveat being that these classic articles (which got a fair bit of attention back in the day and have led to various media appearances and such concerning ‘The Science of Godzilla’) have recently been effectively ruined by the removal of their accompanying images. They’re linked to below.
One more thing. STOP READING NOW IF YOU WANT TO AVOID SPOILERS. THERE WILL BE SPOILERS. Ok, onwards…

Figures in my collection. As a Godzilla fan, it’s great that at least some of these characters have been licensed anew for a modern movie franchise. Image: Darren Naish.
Musical Tunes. The score to this movie – by composer Bear McCreary – was fantastic, and hopefully you noticed that the main monster characters had their own musical themes. These themes incorporated components of the original themes created for the monsters in the original Toho movies. I mean, Godzilla’s theme is from the original 1954 movie and Mothra’s is from the 1962 debut movie, my god. This could partly be considered fan-service but more appropriately reflects an effort to create continuity with these older works. If you liked the music, stay to the end of the credits. There’s a post-credits scenes, by the way, and one that I didn’t predict.

Montage of the main monsters - re-branded as titans for this movie - starring in Godzilla: King of the Monsters. Image: (c) Film Music Central (original here).
Frank Searle’s Nessie. At one point in KOTM, people sift through images when learning about the long cultural influence King Ghidorah has had on human civilizations over the centuries. Among the images we see – ever so briefly – a blurry black and white photo of a water monster, its head obscured because its long, slender neck curves down to the water. This is unmistakeably one of Frank Searle’s infamous Nessie photos. Searle (1921-2005) was a notorious Loch Ness Monster investigator who, between about 1969 and 1983, claimed to have photographed the beast on a great many occasions (over 20; Harrison 1999), often at very close range. As should have been obvious right from the start (amazingly, he was able to dupe some people into thinking that he was the real deal), his photos variously involved models, cleverly posed sticks and branches, and even artwork superimposed onto photos of the water surface. He would likely have been thrilled to see one of his photos appear in a big-budget movie.

One of Frank Searle’s (in)famous Nessie photos. I’m 99% sure that this exact image made a brief appearance in KOTM. Image: (c) Frank Searle.
The Alpha Myth. Main human character Dr Mark Russell is a biologist who’s spent years studying wolves and other animals. In attempting to explain the combative, competitive, hierarchical behaviour observed among the newly emerging titans, he explains that they’re working out which will serve as The Alpha, the big boss animal that gets to control the others. He starts this discussion by saying that animals like wolves work within such a structure, ‘The Alpha’ being the biggest, strongest and meanest and thus the one that gets to rule the pack. This idea is deeply rooted and mainstream in culture and even integral to the philosophy of some animal trainers. But it’s not quite right. Wolf packs are extended families, with the leaders (which can still be called alphas if you want) being the parents of most other pack members. They are thus the most experienced, wisest wolves, not necessarily the baddest or strongest.

Check out all the interesting body language going on in this captive wolf pack. The especially dark animal near the middle is presumably an ‘alpha’. Image: Darren Naish.
I know that this ‘Alpha’ concept was integral to the movie and works fine as an explanation for what we see of titan behaviour. But it would have been nicer if a person who’s supposed to be an experienced biologist said things that better reflected current thinking on the species he was supposed to know best.
The Congolese Titan. At several points during the film, we see world maps which depict titan activity as it’s happening live. And if we look at central Africa during one such scene, we see that the titan stomping around in the Congo region is labelled ‘mokele-mbembe’. Yes, the mokele-mbembe, the long-necked water monster of the Congo, beloved of cryptozoologists and creationists and suggested on many occasions to perhaps be a living sauropod dinosaur. Mokele-mbembe as we ‘know’ it (I mean: as described in the cryptozoology literature) wouldn’t make a particularly impressive titan, since it’s only meant to be about elephant-sized (albeit with a longer neck and tail). The KOTM version is presumably a super-sized version then. I’ve written about mokele-mbembe several times at TetZoo, and also in my book Hunting Monsters (Naish 2017).

One of the most famous of mystery beasts: mokele-mbembe, a creature popularly suggested to be a modern-day amphibious sauropod dinosaur. This illustration is by David Miller for Roy Mackal’s 1987 book on the subject. Image: David Miller/Mackal 1987.
Our Species Name. A reveal in the movie is that palaeobiologist and kaiju scientist Dr Emma Russell has used acoustic data from a mystery species in designing the sounds emitted by the monster-luring ORCA device… aaaand, the mystery species is us, since we’re one of the biggest monsters. What name do they give our species? They have it written – very clearly and in big red letters – ‘Homo sapien’. Major fail, 10 points deducted. It’s difficult to work out why, but many people today seem to think that ‘Homo sapien’ is the correct technical name for our species, and it’s often said this way in TV shows and popular literature. Our scientific name is Homo sapiens, which is scientific knowledge about as advanced as knowing that water has the formula H2O or that the Earth is a sphere.
That’s Not How You Do Scientific Names. On a related note, the titans in the movie have what look like scientific names. But the names don’t make any sense as goes the conventions biologists actually use in naming organisms. I think we’re meant to think that the names we see in KOTM are species names, unique to each kind of titan. These are obviously novel: hypothetically, Godzilla could be something like Gojiratitan terribilis, while Rodan might be, let’s say, Stupendadactylus mexicanus. But KOTM puts all the titans in the same one genus – Titanus (which isn’t available in use since it’s already been used for something else*) – which is then followed by a specific epithet, such that Godzilla is Titanus gojira, Rodan is Titanus rodan and so on. Most viewers won’t care about this, but it’s something that anyone with any knowledge of biology will notice and it’s annoying and a bit dumb. Next time: have an actual biologist on hand to check for these sorts of technical things, they make movies better!
* The longhorn beetle Titanus Audinet-Serville, 1832.

Oh Hollywood, why you no do scientific names right? This image is not from a Godzilla film… Image: (c) Universal Pictures.
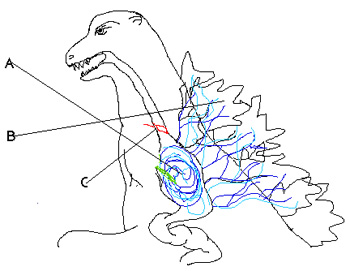
The Hollow Earth. KOTM goes big-time with the idea – previously hinted at in Kong: Skull Island – that our planet is a honeycomb, with hidden tunnels that pass right through it and gargantuan internal chambers and pockets (a description most familiar to modern audiences due to the form of the planet Naboo in Star Wars Episode I: The Phantom Menance). I suppose that this has been worked into the MonsterVerse as a way of explaining how the impossibly huge yet also cryptic titans can remain concealed for a long time and only emerge on occasion.
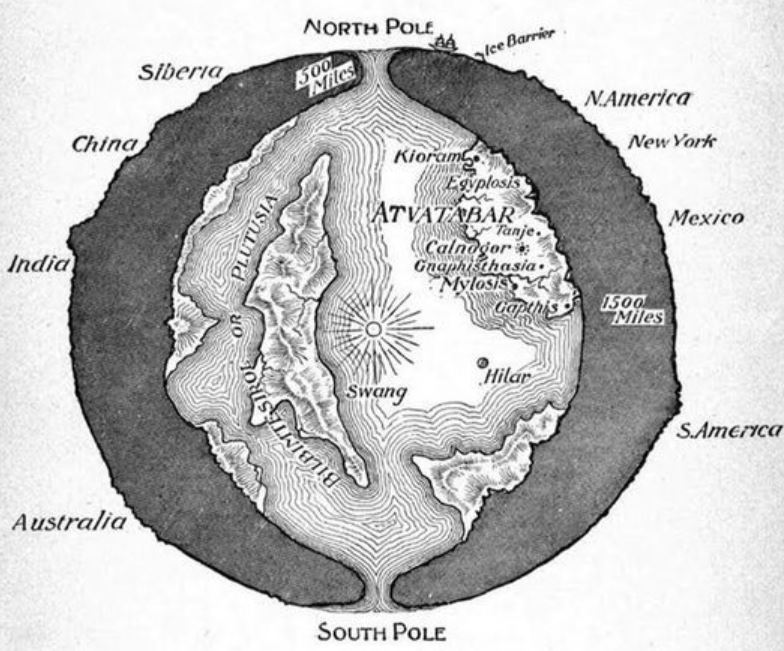
Here’s the ‘Hollow Earth’ image that appears most frequently online (this being because it was uploaded to wikipedia). It’s from William Bradshaw’s 1892 novel The Goddess of Atvatabar. Image: public domain, original here.
The Hollow Earth thing is not unique to the MonsterVerse: it was suggested, as a serious model for our planet’s structure, by Edmond Halley in 1692. Halley didn’t propose the model because it was a fun idea, but because he thought it might explain anomalous compass readings which, he thought, meant that the Earth was formed of more than one rotating sphere. His assumption that compass readings should always be consistent was flawed, since they vary given that the magnetic field is constantly in flux. This model – which involved substantial gas-filled spaces existing between the different sphere – was dead and disproven by the late 1700s, and thereafter it only survived in pseudoscience (UFOs must come from inside the Earth instead of from outer space, and so on) and fiction. Edgar Rice Burroughs is most famous for using the Hollow Earth model for his Pellucidar novels of the early 1900s. These have people tunnelling into the Earth and discovering a hidden world, lit by an inner sun, inhabited by creatures long thought extinct. It’s surprising and interesting to see this idea persist in a modern sci-fi movie series.

The Hollow Earth of Edgar Rice Burroughs is supposed to have looked something like this… [UPDATE: nope, this is nothing to do with the Hollow Earth - it was instead done for the cover of a 1967 magazine issue that celebrated the movie One Million Years BC. Thanks to Alan Friswell for this correction]. This is one of Frank Frazetta’s inimitable illustrations. Image: (c) Frank Frazetta, original here.
That Final Meltdown. Without giving too much away (I know I said that there would be spoilers, but…), the final act of the movie involves Big G discharging A LOT of nuclear energy, so much that he glows red and seems in imminent danger of incandescent eruption. If you’re a hardcore Godzilla fan you may well be predicting at this point that KOTM was going to do its own take on the final scene of the 1995 Godzilla vs Destoroyah (aka Godzilla vs Destroyer) in which Godzilla glows with radiation and eventually (spoiler) melts and dies – only to be replaced by his direct descendant, Godzilla Junior. I don’t know if the KOTM team were inspired by Godzilla vs Destoroyah at all, or if what they did was wholly novel, but the similarities are hard not to notice.

In the 1995 movie Godzilla vs Destoroyah, Godzilla burns up from the inside and endures a painful phase of looking spectacularly patchworked with glowing red. Images: (c) Toho.
Rick and Morty. You might have noticed the prominent Rick Sanchez sticker on the lid of a laptop (if, that is, you’re a Rick and Morty fan. I am). This may or may not be a nod to the fact that Bradley Whitford’s character Dr Rick Stanton was apparently inspired by Rick of Rick and Morty. While I’m here, the original concept of Rick and Morty was based on Doc and Marty of Back to the Future… which is also heavily referenced – or, hell, flat-out mentioned – in another big movie of 2019. A little movie that hasn’t done at all well at the box office.
Finally – Character Continuity. Many of the people in KOTM were previously introduced in 2014’s Godzilla, and therein we got their various backstories (Dr Serizawa, for example, is the descendant of Daisuke Serizawa from the original movie of 1954). But note that they also featured characters who have a direct link to other Godzilla and MonsterVerse movies. The Chen twins – both played by Zhang Ziyi – were an obvious nod to the twin fairies of the 1961 Mothra movie, and Dr Houston Brooks (played in KOTM by Joe Morton, who we mostly know as Dr Myles Bennett Dyson) previously appeared in Kong: Skull Island.

A strength of the new Legendary Godzilla movies is they establish an approximate continuity with the original film of 1954. To go a different route and start Godzilla afresh, I think, is a big mistake. Image: (c) Toho Studios.
And we’ll end this here, I hope you found it interesting. Do you like monsters? Do you like Godzilla? If yes, go and watch this movie. I might go see it again.
For the previous Godzilla-themed TetZoo articles, see…
The science of Godzilla, February 2007
The science of Godzilla, 2010, November 2010
The anatomy of Zilla, the TriStar 'Godzilla', November 2010
Refs - -
Harrison, P. 1999. The Encyclopedia of the Loch Ness Monster. Robert Hale Ltd, London.
Mackal, R. P. 1987. A Living Dinosaur? In Search of Mokele-Mbembe. E. J. Brill, Leiden.
Heilmann, Thompson, Beebe, Tetrapteryx and the Proavian
Many people familiar with ideas on the early evolution of birds and of bird flight will know of the Proavis or proavian, a hypothetical bird ancestor illustrated and discussed by William Pycraft (1868-1942) in 1906 but made better known by Gerhard Heilmann (1859/1861-1946) during the 1910s and 20s. Versions of this creature were later illustrated in colour by Zdeněk Burian for popular books of the 1960s, 70s and 80s. In the article that you’re about to read, we’re predominantly interested in Heilmann’s take on the Proavis.

Caption: fighting, gliding and climbing proavians, depicted in a cluttered woodland environment, by Gerhard Heilmann. This is typical of Heilmann’s many excellent scenes. He used posed plaster models of the creatures before drawing. This illustration was used in the 1916 Danish version of the book (it first appeared in one of his 1912 articles) but does not appear in the 1926 English version. As we’ll see below, the decision to exclude some images from the 1926 version might have served to make it seem more scholarly than its predecessor. Image: Heilmann (1916).
Heilmann was an artist and graphic designer by profession but his interest in bird evolution and anatomy was such that he published a series of articles on the subject for the Danish Ornithological Society between 1912 and 1916 (Nieuwland 2004, Ries 2007).

Caption: self-portrait of Gerhard Heilmann, produced in 1902. Heilmann was an argumentative man often in conflict with “various forms of authority”, and at loggerheads with his own family (Nieuwland 2004). This illustration was included in Nieuwland (2004).
Compiled, these articles described Heilmann’s take on the diversity of ancient fossil birds, embryological development in birds and reptiles, and bird anatomy, with the last in the series explaining what these data meant for the appearance and lifestyle of his Proavis, an imagined animal older and anatomically more archaic than Archaeopteryx. These articles were published together in a Danish-language book Vor Nuvaerende Viden om Fuglenes Afstamning (Heilmann 1916), the title meaning Our Current Knowledge of the Descent of Birds.
Heilmann’s The Origin of Birds. This body of work was sufficiently impressive that – while Heilmann was mostly shunned in Denmark – it was well received elsewhere, and he was encouraged by scientists in Germany in particular to translate it into English. The result was the 1926 The Origin of Birds (republished in the USA in 1927), a scholarly and well-illustrated book that was enthusiastically accepted internationally and almost immediately deemed the most authoritative work on bird origins. The 1926 volume is quite different from that of 1916 in the illustrations it includes (as we’ll see below), but also in how scientific and speculative the text is, some more fanciful sections on palaeobiology being absent from the English version.
It has been said several times that Heilmann’s work was deemed so impressive that, rather than inspire new work on the subject, it effectively bought research on bird origins to a halt. Most relevant workers (albeit not all) now considered the question of bird origins to be resolved. As noted by historian Ilja Nieuwland, things might not have gone this way had the English-speaking readers of Heilmann’s book known that he was an amateur scientist and an artist by trade (Palm 1997, Nieuwland 2004).

Caption: Heilmann’s draftmanship was superb. This illustration (fig. 140 from Heilmann 1926) shows thigh feathering on the chicks of various birds, with 8 being a gliding gecko. “The animal with which we may best compare the bird-ancestor, is the Fringed gecko” [sic] (p. 197). Image: Heilmann (1926).
Heilmann thought that bird ancestors were likely quadrupedal gliders, similar superficially to living gliding geckos (Heilmann 1926, p. 197). This statement is somewhat odd in view of his endorsement elsewhere of a more cursorial view of these animals, but it seems that he imagined them adopting very different poses when on the ground versus when climbing. Heilmann also argued that birds were not dinosaurs – despite his good understanding of the compelling anatomical similarity between theropod dinosaurs and birds – but were instead the descendants of a group closely related to – and supposedly ancestral to – dinosaurs, termed ‘pseudosuchians’*. This was because of his adherence to ‘Dollo’s Law’ – the (erroneous) idea that a lost anatomical structure cannot be regained – and his mistaken contention that dinosaurs lacked clavicles. If dinosaurs couldn’t be ancestral to birds, the true ancestors must have been another group, hence Heilmann’s use of the more archaic ‘pseudosuchians’, already mooted as possible bird ancestors by Robert Broom in his 1913 description of the South African Euparkeria.
* The term pseudosuchian is today applied to the archosaur lineage that includes crocodylians and all of their extinct relatives. Given the historical baggage that comes with the term, I personally don’t think that this is a good idea at all and would prefer it if another name were used for the lineage concerned (like Crurotarsi)… but that’s an issue for another time.

Caption: Euparkeria capensis has long been intimated as a sort of bird ancestor by those looking for such creatures outside of theropod dinosaurs. Not only is this animal only very distantly related to birds, it’s not even part of the crown-archosaur clade. Image: Taenadoman, CC BY-SA 3.0 (original here).
By combining features common to Archaeopteryx as well as to the ‘pseudosuchians’ Aetosaurus, Euparkeria, Ornithosuchus and Saltoposuchus, Heilmann (1916, 1926) described how he invented a creature that looked somewhat like a theropod but had a more archaic skull and foot, retained a fourth metacarpal, and had a smaller pelvis with far shorter pubic and ischial bones.

Caption: Heilmann’s skeletal reconstruction of Proavis, as depicted in the 1916 Danish version of the book… but not in the 1926 English version. Note the lack of long feathers on the hindlimbs. Incidentally, note also that Heilmann was partly responsible for encouraging the belief that forelimb feathers did not cover the hands during the earliest stages of bird evolution. Image: Heilmann (1916).
Heilmann, Thompson and D’Arcy Thompson grids. It’s a matter of some interest that Heilmann didn’t just guess what the imagined morphology of the proavian would be, as might be assumed given the English edition of his book. Instead, he used a technique whereby the relevant anatomical regions were mapped on to a grid and then distorted to mimic the evolutionary process. This grid-based deformation process was pioneered by D’Arcy Wentworth Thompson (1860-1948) and was explained most thoroughly in his 1917 book On Growth and Form (Thompson 1917). It was actually put forward beforehand in 1915 (Thompson 1915).

Caption: Thompson applied his Cartesian grid technique to many animal lineages, his aim being to show that mathematically predictable transformation in one or more anatomical regions could result in profound anatomical change. Here’s one of my favourite examples: how you can derive a molid sunfish (Mola was known to Thompson as Orthagoriscus) from a porcupinefish. Image: Thompson (1917).
By placing grids on top of diagrams of related animals (say, the skull of a human and a chimp), Thompson showed how the skewing of the grid in a certain direction “would result in the sort of changes that would allow the emergence of a new species” (Naish 2017, p. 116). These grids are generally termed D’Arcy Thompson transformation grids, Cartesian transformations or Cartesian grids, and their use was quite popular in the evolutionary literature of the early 20th century. Heilmann was inspired to use the technique after Thompson wrote to him about bird evolution in 1915, and the two wrote to each other on many occasions about the technique and its application to ideas on the evolution of horses, hominids and birds. They also discussed how the resulting illustrations could be used in Thompson’s On Growth and Form (Ries 2007).

Caption: Heilmann included these Cartesian transformations in the 1916 version of his book, but they weren’t included in the 1926 English version. His take on Proavis was not, therefore, simple guesswork. Image: this montage is from Witmer (1991) but the originals are from Heilmann (1916).
Heilmann’s commitment to the technique is demonstrated by the fact that the 1916 Danish edition of his book includes grids that depict the inferred evolution of the avian skull, forelimb and pelvis (I don’t own a copy of the Danish edition, but the diagrams are included in both Witmer (1991) and Ries (2007)). With Ornithosuchus and Euparkeria as ‘starting points’ and Archaeopteryx and modern birds as ‘end points’*, Heilmann used the grids to create relevant intermediates, the results allowing him to generate, piecemeal, his Proavis (Heilmann 1916). A lateral view of the hypothetical animal – showing it in a neural, non-dynamic walking pose – was included in the Danish edition (Heilmann 1916, Ries 2007).
* It should be noted that Heilmann’s Archaeopteryx was not accurate, some of its details (in the skull especially) being semi-hypothetical and informed by Heilmann’s interpretation of ‘pseudosuchians’ like Aetosaurus.

Caption: the more dynamic, climbing version of Heilmann’s Proavis skeletal reconstruction, the only version of the reconstruction included in the 1926 English version of his book. Image: Heilmann (1926).
Remarkably, none of this was included in the English edition at all, this creating the impression that Heilmann simply invented the proavian via intuition and guesswork. He didn’t: it was generated via a testable, repeatable method (albeit using flawed anatomical data). Furthermore, the fact that Heilmann’s skeletal reconstruction of Proavis included in the English edition (Heilmann 1926) is the more dynamic climbing version of the creature gives it a less scientific, more speculative, artistic air than the walking version of 1916.
Heilmann, Beebe and Tetrapteryx. When it came to feathering and other aspects of the integument, Heilmann noted his thoughts on another Proavis-type animal, this time the one invented by Charles William Beebe in 1915. Beebe (1877-1962) was an interesting person. He was a naturalist, ecologist, ornithologist, marine biologist, author and explorer, among other things. Today, he might be best known for the deep-water observations he made while in a bathysphere off the coast of Nonsuch Island off Bermuda during the 1930s.
![William Beebe’s hypothetical tetrapteryx creature. A pre-Archaepteryx glider. You’ll note that Beebe was a pretty good artist [UPDATE: this is an error. The illustrations in this publication were by Dwight Franklin, not by Beebe!]. Image: Beebe (191…](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1546356482204-OC8JSKSSMTH2YL41VQNL/Heilmann-Proavian-Beebe-Tetrapteryx-Jan-2019-Tetrapod-Zoology.jpg)
Caption: William Beebe’s hypothetical tetrapteryx creature. A pre-Archaeopteryx glider. You’ll note that Beebe was a pretty good artist [UPDATE: this is an error. The illustrations in this publication were by Dwight Franklin, not by Beebe!]. Image: Beebe (1915).
When it came to bird origins, Beebe thought that birds originated via a ‘tetrapteryx’ phase where large feathers on the proximal hindlimb were present and functioned in slowing descent during leaping and gliding (Beebe 1915). Both the fossils of Archaeopteryx and the nestlings of living bird species demonstrated the antiquity of these ‘pelvic wings’, Beebe (1915) said. Heilmann (1926) disagreed. His own examination of bird nestlings pertaining to bird lineages across the family tree resulted in his “complete disappointment, for what I found was wholly negative; there was not in any of them the slightest trace of a “pelvic wing”” (p. 194).
As for Archaeopteryx, Heilmann (1926) argued that the hindlimbs were arranged such that they could never have had an aerodynamic function, nor was there evidence for long hindlimb feathering of the sort consistent with ‘pelvic wings’. Ergo, Heilmann illustrated Archaeopteryx without long hindlimb feathering, and he didn’t think that Proavis had them either: his skeletal reconstructions (both the 1916 walking version and 1926 climbing one) lack them (the 1926 climber has short hindlimb feathers, similar in length to those elsewhere on the body) and the gliding proavians included in his various drawings clearly lack them as well. His most famous proavian figure – the semi-erect one standing in a conifer tree (shown below) – does have long feathers on the rear edge of the thigh, though they certainly aren’t as long as they should be as per Beebe’s tetrapteryx model.
The iconic tree-dwelling proavian. The iconic tree-dwelling proavian I have in mind here is one of Heilmann’s best-known and most frequently reproduced illustrations. It has what appears to be great symbolic significance if you’ve read the English version of The Origin of Birds since it’s the very last image in the entire book (Heilmann 1926, p. 199), thus appearing as a visual metaphor for an imagined phrase: “Finally, I can reconstruct the real ancestor of birds, and it looked like this”.

Caption: Heilmann’s most iconic version of his Proavis. Note the long feathers on the back of the thigh and how they’re invisible and thus not deemed important in the gliding individual in the background. Heilmann said that the long feathers close to the base of the tail would produce the required lift in this region. Image: Heilmann (1926).
I was surprised to learn that Beebe produced what looks like his own version of this illustration, and inserted it as a plate in a book published in 1915. However, the image itself is not dated 1915 and does not definitely pre-date Heilmann’s illustration. My suspicion – and that of Paul Stewart, who kindly brought my attention to the image – is that Beebe copied Heilmann’s proavian for his own personal use (the illustration was not intended for publication*). This in itself is interesting since it could mean that Beebe personally endorsed Heilmann’s view of Proavis and regarded it as a ‘descendant’ of his tetrapteryx creature… which it wasn’t, given that Heilmann rejected Beebe’s idea, as we saw earlier. Again, my thanks to Paul Stewart for sharing this image with me and permitting its use here.
* Thanks to Jonathan Kane, I’ve learnt that it was previously published in Tim Berra’s 1977 William Beebe: An Annotated Bibliography.

Caption: William Beebe seems to have produced his own take on Heilmann’s iconic proavian scene. We’re presuming that it was produced some time after Heilmann’s illustration was, but this hasn’t yet been fully confirmed. The illustration appears here courtesy of Paul Stewart.
Today, the significance of Beebe’s tetrapteryx idea is a bit uncertain and, frankly, depends on who you ask. Archaeopteryx probably did have long feathers on the hindlimbs (Longrich 2006), and they perhaps had an aerodynamic role if this animal indulged in aerial locomotion (which it likely did). Furthermore, the discovery of prominent ‘hindlimb wings’ in other feathered dinosaurs (most notably the Chinese dromaeosaur Microraptor) has led some experts to note the predictive power of Beebe’s suggestion (Kane et al. 2016) and to favour the presence and importance of hindlimb feathers in the earliest birds (Longrich 2006, Chatterjee & Templin 2007, Zheng et al. 2013, Xu et al. 2014). However, it’s not at all certain that large, aerodynamically ‘functional’ hindlimb feathers were present consistently across the lineages concerned or that they had the role that Beebe imagined (O’Connor & Chang 2015).

Caption: recent work shows that Archaeopteryx really did have long feathers on the proximal portions of its hindlimbs (these reconstructions are by Longrich (2006)), though they weren’t as long as the feathers Beebe imagined for his tetrapteryx stage creature. Image: Longrich (2006).
As always, there’s more to say and this article is already longer than intended. I also wanted to talk about Pycraft’s initial concept of the Proavis, and also Burian’s depiction, since he didn’t simply copy Heilmann’s illustrations but actually produced yet another hypothetical creature. These things will have to wait to another time. Also worthy of further discussion is the point made earlier about Heilmann’s influence happening despite his status as a ‘mere’ artist, a theme that has parallels elsewhere in the history of vertebrate palaeontology and is relevant to my writings on meme perpetuation, the scientific acceptance of feathering in non-bird dinosaurs and much else besides. I aim to explore these topics (and others) in future articles.
Caption: before Heilmann, there was Pycraft’s proavian of 1906. We’ll have to discuss this creature and its backstory another time. Image: Pycraft (1910).
For previous TetZoo articles relevant to the issues covered here, see…
Flight of the Microraptor, November 2013
The Integrated Maniraptoran, Part 1, May 2016
The Integrated Maniraptoran, Part 2: Meet the Maniraptorans, May 2016
The Integrated Maniraptoran, Part 3: Feathers Did Not Evolve in an Aerodynamic Context, May 2016
Naish and Barrett's Dinosaurs: How They Lived and Evolved, November 2016
Book Review: God's Word or Human Reason? An Inside Perspective on Creationism, March 2018
Refs - -
Beebe, C. W. 1915. A tetrapteryx phase in the ancestry of birds. Zoologica 2, 38-52.
Chatterjee, S. & Templin, R. J. 2007. Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. Proceedings of the National Academy of Sciences 104, 1576-1580.
Heilmann, G. 1916. Vor Nuvaerende Viden om Fuglenes Afstamning. Unknown publisher, Copenhagen.
Heilmann, G. 1926. The Origin of Birds. Witherby, London.
Longrich, N. 2006. Structure and function of hindlimb feathers in Archaeopteryx lithographica. Paleobiology 32, 417-431.
Naish, D. 2017. Evolution in Minutes. Quercus, London.
O’Connor, J. & Chang, H. 2015. Hindlimb feathers in paravians: primarily “wings” or ornaments? Biology Bulletin 42, 616-621.
Palm, S. 1997. The Origin of Flapping Flight in Birds. Svend Plam, Ballerop.
Pycraft, W. P. 1910. A History of Birds. Methuen & Co, London.
Ries, C. J. 2007. Creating the Proavis: bird origins in the art and science of Gerhard Heilmann 1913-1926. Archives of Natural History 34, 1-19.
Thompson, D. W. 1915. Morphology and mathematics. Transactions of the Royal Society of Edinburgh 50, 857-895.
Thompson, D. W. 1917. On Growth and Form. Cambridge University Press, Cambridge.
Witmer, L. M. 1991. Perspectives on avian origins. In Schultze, H.-P. & Trueb, L. (eds) Origins of the Higher Groups of Tetrapods: Controversy and Consensus. Cornel University Press (Ithaca, London), pp. 427-466.
Xu, X., Zhou, Z., Dudley, R., Mackem, S., Chuong, C.-M., Erickson, G. M. & Varricchio, D. J. 2014. An integrative approach to understanding bird origins. Science 346 (6215), 1253293.
Zheng, X., Zhou, Z., Wang, X., Zhang, F., Zhang, X., Wang, Y., Wei, G., Wang, S. & Xu, X. 2013. Hind wings in basal birds and the evolution of leg feathers. Science 339, 1309-1312.
New Living Animals We Want to Find
As a regular denizen of the TetZooniverse, you may well remember the July 2017 article ‘Fossils We Want to Find’ in which I discussed a list of hypothetical fossil things that we might one day discover but haven’t yet. Wouldn’t it be fun to do the same sort of thing with extant species; that is, with discoveries pertaining to living, breathing animals? Over at the Zoology for Enthusiasts facebook group (a spinoff of the Tetrapod Zoology facebook group), Jordan Fryer suggested doing exactly this, and consequently people have been coming up with their own suggested living animals that might await discovery. Because this seemed like a lot of fun (and a chance to discuss some really neat and unusual stuff), I thought I’d give it a go.

Caption: the 2017 precursor to the article you’re reading here was all about fossil animals. It included this photo, which shows me in the act of discovering a dinosaur bone in the Moroccan Sahara. Image: Richard Hing.
Naturally, any list of this sort is horribly subjective, reflecting the interests and biases of the person compiling the list, but so be it. It also seems all too easy to turn any compilation into a ‘list of most discoverable cryptids’: for those of you who don’t know, I have a long-standing interest in cryptozoology and have published on it quite frequently (see Naish (2017) for starters). For the most part, I’ve not done this, though read on.

Caption: many of my thoughts on mystery animals can be found in my 2017 book Hunting Monsters. I am not - sorry - much impressed by the case for such supposed animals as the mokele-mbembe, an artistic reconstruction of which is shown at right. Image: David Miller, in Mackal (1987).
I’ve also mostly excluded hypothetical discoveries that are inspired by the creatures of cryptozoology but could arguably be considered independent of the cryptozoological literature. In part this is because I don’t think they’re plausible or worth considering, but it’s also because they’re cliched and the opposite of original. So, no ‘living sauropods from the Congo’ or ‘living plesiosaurs in Loch Ness’, for example.
As for what I have selected: well, some of my suggestions are sillier than others, and some are perhaps not that interesting to non-specialists. But, whatever. Feel free to dissect my suggestions in the comments, and perhaps come up with your own.

Caption: among my suggested ‘fossils we want to find’ are protobats (like the hypothetical examples shown at left, from Graham (2002)) and a good skeleton of the giant hominid Gigantopithecus blacki. This ilustration of a lower jaw is from Simons & Ettel's (1970) magazine article. Images: Graham (2002), Simons & Ettel (1970).
A habitually bipedal, large, non-human hominid. Whatever you think of all those stories, anecdotes and sightings about bigfoot, yeti, almas, orang-pendek, yowie and so on and on, the fact remains that the discovery of a large, bipedal non-human hominid – whether it be a pongine, hominine, or member of another hominid lineage – would be a huge deal. It would not just be one of the most newsworthy creatures to ever be discovered; it would also have enormous ramifications for our understanding of hominid evolution and potentially the human condition itself.

Caption: are crypto-hominids a cultural phenomenon more than a zoological one? I’ve argued for both possibilities at different times. Whatever… for the purposes of the article you’re reading now, I hope we can agree that the discovery of such an animal would be high on any hypothetical ‘wants’ list. Image: Darren Naish.
It would also – if relating to North America or northern Eurasia in particular – very likely have a significant impact on economy, land management and land use in those regions… or, you’d hope it would, anyway (who knows, given the current state of environmental protection in the USA). The hypothetical discovery of such an animal would also be regarded by many as one of the biggest ‘wins’ ever scored against ‘establishment science’, and thus could well be a bad thing (viz, “if scientists were wrong about this, what else could they be wrong about?”). And I’ll stop there before we dive into a rabbit-hole of conspiracy theories and coverups.
A big, flightless passerine. The majority of living bird species – over 60% of them – are passerines, or perching birds. This is the great group that includes crows, thrushes, warblers, finches, sparrows and so many others. For all their success, wide distribution and diversity, passerines are generally quite samey. There are no big, long-legged wading passerines, or heavy-bodied diving passerines or flightless running passerines, for example. Why this is so remains mysterious: passerines didn’t take to those niches because… well, they just didn’t. Does this mean that they couldn’t? As usual, we can come up with a few reasons as to why they were ‘constrained’ in evolutionary potential, but any one of those reasons could be overturned by some evolutionary deviant that refuses to pay attention to the rules.

Caption: passerine birds are diverse, to a degree… here’s just a sample of their diversity. This is part of a giant montage that’s being built for my in-prep textbook The Vertebrate Fossil Record. Image: Darren Naish.
And thus I submit that a particularly large, wholly flightless, cursorial passerine should make itself known to the world. It should be a record-holder as goes size, but not necessarily be that much bigger than the largest known passerines (like lyrebirds and ravens): I’m talking about a bird that weighs 3-5 kg and is thus similar in size to a large chicken. It should be a big, long-legged rail-babbler, quail-thrush or similar, and hence be a denizen of Wallacea or nearby.

Caption: Eupetes, the Malaysian rail-babbler. A hypothetical big, flightless passerine should be a close relative of this bird. Image: Francesco Verronesi, CC BY-SA 2.0 (original here).
A few recently extinct, island-dwelling passerines were flightless, so we do know that passerines have the evolutionary potential to follow this pathway. Such species (a bunting and a few New Zealand wrens… and possibly a few others) were all small (less than 40 g).
A western Asian giant salamander. Giant salamanders (cryptobranchids) are restricted today to eastern Asia (where Andrias occurs) and North America (where Cryptobranchus occurs). Hunting, human disturbance, habitat loss and deterioration, climate change and other issues are putting them into perilous decline, right at the same time as we’re discovering that some of them are species complexes. They were more widespread in the past than they are today, since fossils show that Andrias salamanders were widespread across Europe and Asia between about 28 and 2 million years ago.

Caption: an Asian giant salamander (Andrias) photographed in captivity. Record-holding specimens of Andrias can be 1.8 m long and exceed 60 kg, and some extinct species reached even larger sizes. Image: Markus Bühler.
While there are very good reasons for the decline and extinction of the animals in the areas concerned, some of the regions where they formerly occurred still have what look like suitable habitat today and are sparsely populated by people. Furthermore, extinct giant salamanders weren’t all denizens of fast-flowing, highly oxygenated streams like those inhabited by the modern populations. Some inhabited ponds and lakes. Ergo: I would really, really like there to be a west Asian cryptobranchid that comes from a habitat considered weird for the other living members of the group. And it doesn’t have to be a giant of 2 metres or more. A hellbender-sized species of 70 cm or so will do fine thank you very much.

Caption: some extinct cryptobranchids - this is Zdeněk Burian’s reconstruction of Andrias scheuchzeri - inhabited European ponds and lakes. I’ve previously criticised this image for showing the animal as terrestrial. Since then, the proposal has been made that some extinct cryptobranchids (albeit not A. scheuchzeri) were significantly more terrestrial than living species. Image: (c) Zdeněk Burian.
Again, this is an area of special interest to cryptozoologists, since there have been occasional suggestions that stories, engravings and such from western Asia might reflect folk knowledge of unusually big salamanders in the region. In reality, the images and stories concerned are super-ambiguous and more likely refer to otters and god knows what else.
Caption: at left, an Andrias skull. Image: Darren Naish. At right: Japanese giant salamander (A. japonicus) illustration by Y. de Hoev from 1887. Image: Y. de Hoev, public domain (original here).
A living albanerpetontid. Everyone knows that there are three main groups of living amphibian: caecilians, salamanders and anurans (frogs and toads). But until (geologically) recently, there was a fourth group: the albanerpetontids, sometimes termed albies by those who work on them. Albanerpetontids were geographically widespread, their range including Eurasia, northern Africa and North America, and they were geologically long-lived too. The oldest are from the Middle Jurassic while the youngest are… well, we’ve known of Miocene fossil albanerpetontids for decades, have known of Pliocene specimens since 2005 (Venczel & Gardner 2005), and now know that at least one species persisted into the Pleistocene (Villa et al. 2018). The fact that their fossil record has been creeping towards the Recent means that the possibility of fossil and even extant Holocene specimens being discovered isn’t ridiculous, especially given the small size of these animals and hence tiny size of their bones.
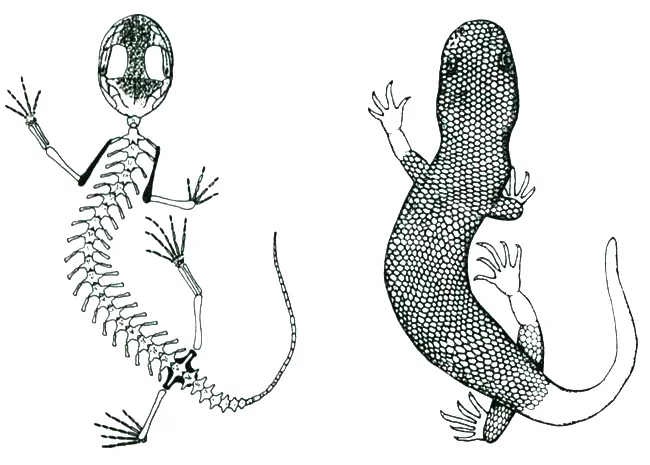
Caption: new salamander species are occasionally discovered in Europe and Asia even now. It would be amazing if an animal suspected to be a ‘new salamander’ one day turned out to be a living albanerpetontid. These reconstructions were published by McGowan & Evans (1995). They might have erred in implying that the scales would be externally visible as shown here; more likely is that they were concealed by epidermis, as in other scaly fossil amphibians. Image: McGowan & Evans (1995).
To be frank, a live albanerpetontid wouldn’t be a particularly spectacular animal: it would be a tiny, slim, salamander-like amphibian less than 10 cm long, and it wouldn’t be much fun to watch since it would spend most of its time hiding and burrowing in leaf litter. But among herp-nerds it would be a huge deal. Live albanerpetontids were scaly-skinned (though the scales were not necessarily visible externally), with eyelids, and with adaptations in the snout, skull-roof, neck and body shape linked to head-first burrowing (McGowan & Evans 1995).

Caption: an artistic reconstruction of a live albanerpetontid… produced for my in-prep The Vertebrate Fossil Record. Image: Darren Naish.
A Eurasian palaeognath. Palaeognaths are the big, flightless ratites (ostriches, emus and so on), the superficially gamebird-like, flight-capable tinamous, and their extinct relatives. A huge amount has been written about the evolutionary history and biogeography of these birds, since their distribution is curious and has resulted in all kinds of different models about how they might have spread around the world. I’ve written about this issue at length on previous occasion (the articles concerned being famous for generating the longest-ever comment threads in the history of TetZoo… though all of this is mostly wasted now, what with SciAm’s paywalling of the site, sigh). Living palaeognaths are absent from Eurasia, despite the former present in the region of ancient, flight-capable Paleogene taxa, extinct ostriches and others.

Caption: as this map shows, modern palaeognaths occurred everywhere until recently (except Antarctica) with the exception of northern North America and the cooler parts of Eurasia. Extinctions across Eurasia, Madagascar and New Zealand of course saw the disappearance of various members of the group. Image: Darren Naish.
The fact that Paleogene Europe was home to many bird groups that no longer occur there but are now denizens of tropical regions elsewhere leads me to hope for a living palaeognath – a tinamou- or bustard-sized species – that descends directly from archaic Paleogene taxa and now lives in the Asian tropics. It should be a cryptic generalist with barred plumage and a mid-length bill and a reduced flight ability.
A gigantic, predatory, limbed amphisbaenian. Regular readers of TetZoo might know that I really like amphisbaenians: the mostly limbless, bullet-headed ‘worm lizards’ of the American tropics, Africa, and parts of southern Europe and western Asia. Amphisbaenian evolutionary history and biogeography has become increasingly complex in recent years as we’ve learnt a bunch of new stuff about their fossil history, genetics and anatomy. Among the weirdest of amphisbaenians are the ajolotes (or bipedids), the only extant group to possess limbs. These limbs are not small stumps or flaps (as they are in some other near-limbless, serpentine squamates) but well-developed, clawed forelimbs. According to some phylogenetic models, ajolotes are not the sister-group to limbless amphisbaenians but deeply nested within the limbless clade (Conrad 2008, Videl et al. 2008), in which case their limbedness – if you will – perhaps evolved from limbless ancestors. Add to this the fact that some amphisbaenians are robust-jawed, short-faced predators of vertebrates that ambush prey from beneath the surface and bite chunks from the bodies of surface-dwelling mammals and reptiles.
Caption: Bipes, an ajolote of Mexico (they might occur in parts of the USA as well). Three extant species are recognised. Image: Darren Naish.
So then… where oh where are the giant, limbed, robust-skulled, vertebrate-eating amphisbaenians? By ‘giant’, I am not talking about a graboid-sized monster of several metres (though that would be nice), but a more reasonable animal of a mere 1.5 metres or so. Easily the stuff of nightmares. They could inhabit warm regions of any continent.

Caption: Carl Gans’s illustration of a burrowing ajolote, showing how the large, well-clawed forelimbs function in propulsion. This is clearly a Five-toed worm lizard Bipes biporus; the other extant species have four and three digits, respectively. Image: Gans (1974).
Is there any reason to think that gigantic, predatory, limbed amphisbaenians might actually exist and await discovery? Nope. But I wish it were so. Regular readers might recognise that such creatures are denizens of the alternative-timeline Earth of the Squamozoic, but I’m sure that that’s coincidental.
Caption: what would a gigantic, predatory, limbed amphisbaenian look like? Like this, of course. Image: Darren Naish.
An African or west Eurasian, long-beaked river dolphin. On several occasions within the history of odontocete cetaceans (‘toothed whales’), lineages have moved into brackish and estuarine environments, and eventually made the transition to committed freshwater life. There are the Asian Platanista species, the recently extinct Lipotes of China, and the tropical American Inia species. Once united within Platanistoidea and thought to be close kin, we know today that these animals represent at least three separate transitions to the freshwater environment (the term Platanistoidea is now restricted to the Platanista lineage alone). In addition, members of other groups – I’m thinking of the delphinid Orcaella – occur in rivers within parts of their range. There’s also a fossil beaked whale that might be indicative of freshwater specialisation in yet another odontocete group (Mead 1975).

Caption: river dolphins are pretty special looking. This is a Ganges river dolphin (Platanista gangetica). Image: Zahangir Alom / Marine Mammal Commission / National Oceanic and Atmospheric Administration, public domain (original here).
In view of all this, why aren’t there river-dwelling dolphins in Africa, Europe or western Asia? Again, the answer seems to be… there just aren’t. A few fossil taxa suggest that such animals might have evolved if things had gone another way (there are fossil platanistoids from the Caucasus, for example). But I humbly submit that the great river systems of tropical Africa, the Tigris-Euphrates system of western Asia and the Danube, Po, Ebro, Dniester and others of southern Europe would be much improved if only we knew of their endemic riverine dolphins. I’m talking about a true riverine specialist, convergent with Inia and Platanista, with a long beak, spike-like teeth, reduced eyesight, the works. And if you want to play fast and loose with antiquarian literature and anecdote, there are references in the literature to ‘river dolphins’ in the Nile and there are even one or two eyewitness accounts from central Europe that describe long-beaked ‘dolphins’ seen in rivers and lakes.

Caption: if there are extant west Eurasian or African river dolphins, they should look like this. This is a hypothetical species, modelled on the American Inia and Asian Platanista. Image: Darren Naish.
An endoparasitic tetrapod. Tetrapods have become parasites on several occasions. Vampire bats are parasites of birds and mammals, and it’s even been argued that some blood- and milk-eating human populations can be considered parasites of the mammals they rely on (though the mammals concerned are domesticated, so it’s complicated). Elsewhere among vertebrates, everyone knows about the parasitic catfishes that invade the gills of other actinopterygian fishes and even the urethras of mammals; less familiar is the fact that other actinopterygians can, on rare occasion, become trapped inside the bodies of other vertebrates and then make a successful living. Yes, you read that right. I have in mind the case where two Snubnosed or Pugnose eels Simenchelys parasitica were discovered living inside the heart of a mako shark (Caira et al. 1997; see also Eagderi et al. 2016). This eel is not – despite its name* – ordinarily an internal parasite: this was a case of facultative endoparasitism!

Caption: at left, a snubnosed eel found living inside the heart of a shark. Eels are not tetrapods, it’s true. But here’s evidence that aquatic vertebrates can become endoparasites. Image: Caira et al. (1997). At right: an aquatic typhlonectid caecilian. Surely it’s only a matter of time before we discover an endoparasitic one of those as well. Image: Neil Phillips.
There are all kinds of reasons why a tetrapod couldn’t become an endoparasite, respiration being high on the list. A hypothetical endoparasitic tetrapod would have to be small, with remarkable tolerance of unusual chemical and thermal conditions, with low oxygen requirements, and most likely with the ability to respire cutaneously or via gills. In other words, it should be the world’s weirdest caecilian. As if caecilians aren’t weird enough, I’d love there to be small, endoparasitic caecilians. Given that some caecilians are already aquatic gill-breathers that will consume the tissues of fish (exhibit A: the sequence from River Monsters where Jeremy Wade discovers swarming typhlonectid caecilians in the carcass of a large fish), I predict these animals to be aquatic, South American species that parasitise actinopterygians and aquatic mammals, like Inia the river dolphin.
* Snubnosed eels were given the name ‘parasitica’ because they opportunistically latch on to the bodies of larger fish and eat away at the flesh. They were not thought to ever be proper internal parasites prior to 1992.
And that’s where I’ll stop for now. I actually came up with a list containing numerous additional ‘wish list’ animals but time is against me. Maybe I’ll cover them in another article. Whatever, this was all a bit of fun and I hope you enjoyed it.
For TetZoo articles relevant to the issues covered here, see…
Surreal caecilians part I: tentacles and protrusible eyes, January 2008
Surreal caecilians part II: pass mum’s skin, hold the mayo, January 2008
Close up to Andrias, despite the smell and the teeth, December 2010
Welcome to the Squamozoic!, April 2013
If Bigfoot Were Real, June 2016
PS I’m going to stop linking to the SciAm run of TetZoo articles soon, because I cannot access them at all and they’re now all but useless. They all need to be relocated to an open-access site.
Refs - -
Caira, J. N., Benz, G. W., Borucinska, J. & Kohler, N. E. 1997. Pugnose eels, Simenchelys parasiticus (Synaphobranchidae) from the heart of a shortfin mako, Isurus oxyrinchus (Lamnidae). Environmental Biology of Fishes 49, 139-144.
Conrad, J. 2008. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bulletin of the American Museum of Natural History 310, 1-182.
Eagderi, S., Christiaens, J., Boone, M., Jacobs, P. & Adriaens, D. 2016 Functional morphology of the feeding apparatus in Simenchelys parasitica (Simenchelyinae: Synaphobranchidae), an alleged parasitic eel. Copeia 104, 421-439.
Graham, G. L. 2002. Bats of the World. St. Martin’s Press, New York.
Mackal, R. P. 1987. A Living Dinosaur? In Search of Mokele-Mbembe. E. J. Brill, Leiden.
McGowan, G. J. & Evans, S. E. 1995. Albanerpetontid amphibians from the Cretaceous of Spain. Nature 373, 143-145.
Mead, J. G. 1975. A fossil beaked whale (Cetacea: Ziphiidae) from the Miocene of Kenya. Journal of Paleontology 49, 745-751.
Naish, D. 2017. Hunting Monsters: Cryptozoology and the Reality Behind the Myths. Arcturus, London.
Simons, E. L. & Ettel, P. C. 1970. Gigantopithecus. Scientific American 222 (1), 77-84.
Venczel, M. & Gardner, J. D. 2005. The geologically youngest albanerpetontid amphibian, from the Lower Pliocene of Hungary. Palaeontology 48, 1273-1300.
Vidal, N., Azvolinsky, A., Cruaud, C. & Hedges, S. B. 2008. Origin of tropical American burrowing reptiles by transatlantic rafting. Biology Letters 4, 115-118.
Villa, A., Blain, H.-A. & Delfino, M. 2018. The Early Pleistocene herpetofauna of Rivoli Veronese (Northern Italy) as evidence for humid and forested glacial phases in the Gelasian of Southern Alps. Palaeogeography, Palaeoclimatology, Palaeoecology 490, 393-403.
The Dougal Dixon After Man Event of September 2018
“Speculative biology, or speculative evolution, is a term that refers to a very hypothetical field of science that makes predictions and hypotheses on the evolution of life in a wide variety of scenarios and is also a form of fiction to an extent. It uses scientific principles and laws and applies them to a "what if" question” — the Speculative Evolution Wiki

Caption: Dixon 1981; Dixon 2018.
On September 11th 2018, I had the extraordinary privilege of appearing on stage with author, artist, editor, model-maker and visionary Dougal Dixon to discuss his famous book of 1981 After Man: A Zoology of the Future (Dixon 1981). We were joined by more than 215 interested members of the public, effectively filling the venue (Conway Hall in London).

Caption: at Conway Hall once again, such a noble venue. Image: Will Naish.
As you might have gathered if you’re a regular Tet Zoo reader, After Man has just been republished, and our on-stage event – hosted by New Lands London, and arranged by Scott Wood – also served as the launch for the new, 2018 edition (Dixon 2018). This was on sale and available for signing at the meeting.
As you can see from these photos, Dougal brought along a treasure trove of material relevant to the genesis of After Man, including his original sketches, text, draft spreads and the original page plan for the entire work. The latter consisted of large card mock-ups with small, rectangular vignettes depicting the planned look for each page. Dougal explained how he took these documents along to two – yes, just two – publishers and immediately got the green light from both. Evidently, he knew exactly what he was doing.

Caption: Dougal on stage, showing the original page plan for After Man. Small vignettes, showing the planned look of all the pages, are arranged in sequence. Image: Darren Naish.
The original sketches are excellent and a testament to Dougal’s skill and planning. The quality of these illustrations also leave you wondering why it isn’t Dougal’s art that we see in the final book, and I impressed upon him during our discussion how fantastic it would be to one day see these unpublished illustrations in another book: a ‘The Making of After Man’ or something along those lines. We’ll see.

Caption: foreign language translations of After Man, a Vortex and Raboon model, and relevant magazine issues (like the October 1981 ish of BBC Wildlife). (c) Dougal Dixon. Image: Darren Naish.
Several of the creatures pictured on the draft spreads were obvious prototypes of versions that made it into the final book but others were evidently abandoned at some point. A few of those ‘prototypes’ showed how the original animals had a different look relative to the published descendants: the bone-cracking Ghole Pallidogale nudicollum, to take one example, looked a lot more like a big mongoose in its original guise than is obvious in the book.

Caption: original text and original draft double-page spread for After Man, showing creatures inhabiting tropical grasslands. You’ll recognise some (but not all!) of the creatures as the prototypes of versions that made it to final publication. (c) Dougal Dixon. Image: Darren Naish.
Dougal also brought foreign-language editions of After Man and a few models of After Man’s creatures with him, including a resin model of the Vortex Balenornis vivipara (a reasonable number were made, but it may be that this is the only one still in existence) and a wonderfully detailed Desert leaper Aquator adepsicautus. Alas, the Night stalker Manambulus perhorridus model pictured on the dustjacket of Dixon 1981 – the model I most wanted to see – is not in Dougal’s possession so was a no-show.

Caption: Desert leaper model. For the handful of you that haven’t read After Man, the Desert leaper is a giant, desert-dwelling muroid rodent (in cases more than 3 m long) that undergoes significant fluctuation in fat deposition (and hence mass) according to season. (c) Dougal Dixon. Image: Darren Naish.
As goes our actual discussion, we covered the backstory to After Man (some of which will be familiar if you know the interview I published at ver 3 back in 2014), the response from critics and reviewers and the many overseas trips Dougal got to enjoy as a consequence of the book’s success, the substantial interest from Japanese markets and the Japanese stop-motion and animated movies (we watched a short segment from the stop-motion movie, copies of which were given away on DVD to people buying the book), the various efforts by studios in Hollywood to get an After Man movie off the ground, and the connection between After Man and Dougal’s more recent project Green World (thus far only published in Japanese).

Caption: original sketches, by Dougal, of creatures illustrated for After Man. The animals were then re-illustrated by various other artists. (c) Dougal Dixon. Image: Darren Naish.
We finished with a Q&A session and audience participation. Questions included the ‘new look’ Night stalker (yup… I shall say no more), Dougal’s thoughts on the future of humanity, how and which fossil species had influenced the creatures of After Man, and what might be different in After Man if Dougal were to write the book today.

Caption: the Vortex model that Dougal brought along. It’s about 60 cm long. (c) Dougal Dixon. Image: Darren Naish.
For a lifelong fan of After Man and Dougal’s connected writings, this event was an absolute thrill and I’m tremendously happy to have been involved. And judging by our audience’s response, it was enjoyed by everyone who attended too: thanks so much to everyone who came along and participated.
Two final things are worth saying. Firstly, I was asked innumerable times whether the event was going to be recorded. Alas, I was simply unable to organise this or even remember it given all the other stuff I had to worry about, though I think (and hope) than an audio recording exists. Secondly, this is not the only Dougal Dixon-themed event of 2018! Dougal is also on stage at this year’s TetZooCon when he will be joined by Gert van Dijk (of Furahan Biology and Allied Matters) in a discussion on speculative biology. TetZooCon happens on Oct 6th and 7th in London and tickets are still available. Dougal will also be bringing archive material to that meeting as well!

Caption: Darren Naish (l) and Dougal Dixon (r) on stage at Conway Hall, September 2018. Image: Will Naish.
My thanks to Dougal for being such a brilliant person to talk to and for all the material he brought along, to Scott and everyone else at Conway Hall and New Lands for organising things and setting it all up, to the Breakdown Press people for the book selling, and to our brilliant audience for their interest, enthusiasm and participation.
For previous articles on speculative biology, see…
Speculative Zoology: Wedel throws down the gauntlet, February 2007
Oh no, not another giant predatory flightless bat from the future, March 2007
Come back Lank, (nearly) all is forgiven, September 2008
The Tet Zoo guide to the creatures of Avatar, January 2010
The science of Godzilla, 2010, November 2010
The anatomy of Zilla, the TriStar ‘Godzilla’, November 2010
Giant flightless bats from the future, November 2010
Welcome to the Squamozoic!, April 2013
Of After Man, The New Dinosaurs and Greenworld: an interview with Dougal Dixon, April 2014
The LonCon3 Speculative Biology event, August 2014
Summer 2018 at Tet Zoo Towers, July 2018
Speculative Zoology, a Discussion, July 2018
Could We Domesticate (Non-Bird) Dinosaurs?, August 2018
Refs - -
Dixon, D. 1981. After Man: A Zoology of the Future. Granada, London.
Dixon, D. 2018. After Man: A Zoology of the Future. Breakdown Press, London.



